
青海高原鼠疫菌DFR分型及空间分布特征的研究
2014-04-09 03:18:24杨晓艳魏柏青辛有全熊浩明魏荣杰祁美英李存香代瑞霞
中国人兽共患病学报 2014年4期
杨晓艳,魏柏青,辛有全,靳 娟,何 建,熊浩明,魏荣杰,祁美英,李存香,金 泳,代瑞霞
青海省自1954年证实存在鼠疫自然疫源地以来,全省有33个县(市)、119个乡(镇)被确定为鼠疫疫源地,疫点695处,疫源面积近20万km2。青海高原鼠疫自然疫源地是目前我国动物间和人间鼠疫流行最严重的地区之一,至2012年的60年间,仅1984年、2007年、2008年和2012年无病例报告外,其余年间均有人间鼠疫发生。因此,迫切需要采用新的技术手段,针对这种具有重要公共卫生意义的传染病进行深入研究,掌握其遗传变异的规律、形成机理以及地域和时空分布特征,为制订有效的防治策略,鼠疫菌的分型、溯源以及保障国家生物安全奠定基础。为此,作者对青海高原1954年以来分离的839株鼠疫菌采用DFR技术对其进行基因分型的研究。
1 材料与方法
1.1实验菌株 共有839株鼠疫菌用于本研究。其中喜马拉雅旱獭鼠疫自然疫源地829株,青海田鼠鼠疫自然疫源地10株。这些菌株涵盖了青海高原鼠疫疫源地的不同时间、地区、宿主、媒介体内分离的鼠疫菌,由青海国家鼠疫菌保藏中心提供。
1.2鼠疫菌DNA的提取 将鼠疫菌培养于赫氏琼脂培养基上,28 ℃培养24 h,DNA提取按照经典的苯酚-氯仿混合抽提法进行[1]。
1.3引物和PCR扩增方法 鼠疫菌差异区段(Different Region,DFR)分型,使用23个DFR和PMT1(质粒验证引物)的扩增引物(引物序列参见文献[2])。PCR反应体系:10×buffer 3 μL,10 mmol dNTP 0.1 μL, 5U/μL Taq DNA聚合酶0.2 μL, 10 μmoL引物对1 μL,模板DNA(2 ng/μL)5 μL, 用去离子水补足至30 μL。PCR扩增条件:95 ℃预变性3 min,95 ℃变性40 s,56 ℃退火40 s,72 ℃延伸40 s共进行30个循环,最后72 ℃终延伸3 min。PCR产物检测:扩增结束后取10 μL产物进行浓度为2%的琼脂糖凝胶电泳,用goldview 染料进行染色,紫外灯下记录结果,阳性结果记录为“1”,阴性结果记录为“0”。每次PCR检测均设阴性对照(用去离子水取代模板)和阳性对照(模板为鼠疫菌菌株82009和620024 DNA的混合物,包括24个DFRs)。可疑PCR结果和阴性PCR结果至少重复1次。
1.4地理信息系统(GIS) 运用青海省地方病预防控制所地理信息系统(GIS)[3]进行鼠疫菌基因型空间分布特征的分析。
2 结 果
2.1青海省鼠疫菌的基因分型 青海高原存在2种类型的鼠疫疫源地。被试的839株菌,共发现有11个基因组型。检索包含909株中国鼠疫菌自然分离菌株的DFR分型数据库[2],有8个基因组型与其相同,分别为1b、5、7、8、10、14、21、30型等基因组型,同时我们还发现了3个新的基因组型,即32、44、36。32型与5型接近,但32型缺失了DFR18;44型与8型接近,44型也缺失DFR18;36型亦与5型接近,但与5型相比缺少DFR10和DFR18。青海高原喜马拉雅旱獭鼠疫疫源地以5、8型为主,青海高原青海田鼠鼠疫疫源地鼠疫菌基因组型为14型。青海省鼠疫菌株23个DFR和PMT1(质粒验证引物)分布见表1。

表1 24个DFR在青海省791株鼠疫菌中的分布状态
Note: “1” means PCR amplification showed positive; “0”means PCR amplification showed negative.
2.2鼠疫菌基因型的空间分布特征 青海省鼠疫疫源地所分离的鼠疫菌基因型具有一定的地理分布特征,8型菌株的分布区为祁连山南麓、青海湖环湖地区、青海南山的祁连、门源、刚察、海晏、共和、天峻、乌兰和宗务隆山的德令哈等地区,占56.14%(471/839);5型菌株分布于青南高原的玉树、杂多、治多、称多、囊谦、曲麻莱和海西西部的格尔木等地区,占23.12%(194/839);1b型菌株分布于冷湖、乌兰、泽库;7型菌株分布于循化、同仁、泽库、德令哈、囊谦;32型菌株分布于格尔木的唐古拉地区;44型菌株分布于祁连、门源;36型菌株分布于囊谦;唯独囊谦地区有10型分布,其菌株分离自2004年一次肺鼠疫流行中的尸体和病人。在称多县青海田鼠分布区,鼠疫菌基因型属于14型,占1.19%(10/842)。见表2。
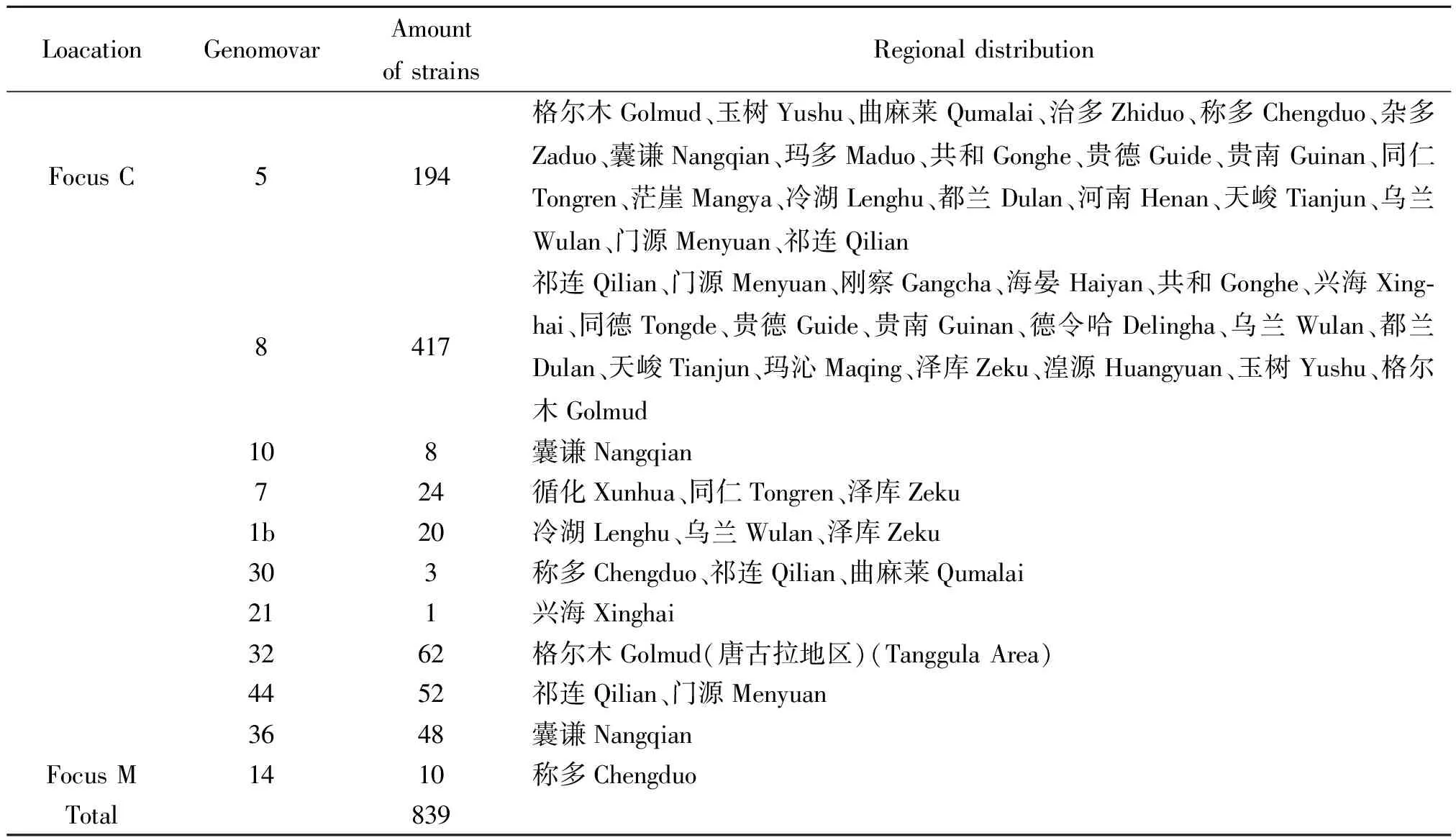
表2 青海高原鼠疫耶尔森菌基因组型的地区分布
Note:Focus C--Marmothimalayanplague foci; Focus M--Microtusfuscusplague foci
3 讨 论
周冬生等[4]人利用全基因组芯片比较基因组学的研究获得了鼠疫菌22 个差异区段(DFR),并将我国各鼠疫自然疫源地分离的 260 株鼠疫菌分成14 个基因组型。之后,军事医学科学院微生物流行病研究所又通过消减杂交技术发现了新的差异区段,并使用23个DFR的扩增引物,对分离自我国的909株鼠疫菌进行了DFR分型,将909株中国鼠疫菌分离株分成32个基因组型,根据疫源地与宿主的分布规律,提出了主要基因组型和次要基因组型的概念,前者与鼠疫菌在自然疫源地中的演化密切相关。因此,随着尽可能多的DFR的发现,DFR分型方法会日趋完善。
青海省青海田鼠鼠疫自然疫源地是近年来新发现的一块疫源地[5],青海田鼠是主要储存宿主,细钩黄鼠蚤和直缘双蚤指名亚种为主要传播媒介。分布于青海省称多县珍秦乡境内,与四川省石渠县接壤。截止目前青海田鼠鼠疫疫源地内分离到的10株鼠疫菌均为14型。
青海省喜马拉雅旱獭鼠疫疫源地内鼠疫菌基因型比较复杂,以5、8型为主。5型多分布在青海的青南高原和海西西部地区; 8型菌株的分布区为祁连山南北麓、青海湖环湖地区及青海南山和宗务隆山等地区;8型和5型的交汇处常同时有2~3种基因型的流行; 2004年青海省囊谦县肺鼠疫暴发流行时所分离的8株菌株其基因组型全部为10型。新发现的3个基因组型,32型菌株分布于格尔木的唐古拉地区;44型菌株分布于祁连、门源;36型菌株分布于囊谦。由于青海高原地形复杂,自然景观垂直成带十分明显,景观特征的多样性使得地区间生态系差异较大,草甸草原几乎占据整个青海;森林草甸草原出现于祁连山东部、玉树东南部;昆仑山—阿尔金山山地以干旱的高山草原为主,鼠疫菌为了适应不同的自然景观,基因组型发生变化也就不可避免。
参考文献:
[1]Achtman M, Zurth K, Morelli G, et al.Yersiniapestis, the cause of plague, is a recently emerged clone ofYersiniapseudotuberculosis[J]. Proc Natl Acad Sci U S A, 1999, 96(24): 14043-14048.
[2]Li Y, Dai E, Cui Y, et al. Different region analysis for genotypingYersiniapestisisolates from China[J]. PLoS ONE, 2008, 3(5): e2166. DOI:10.1371/journal.pone.0002166
[3]Tang XY, Wang GJ, Li Y, et al. The establishment of multi-diseases geographic information system for the endemic diseases in Qinghai province[J]. Chin J Endemiol, 2010, 29(6): 687-689. (in Chinese)
唐新元,王国钧,李勇,等.青海省地方病多病种综合地理信息系统的建立[J].中国地方病学杂志,2010,29(6):687-689.
[4]Zhou DS, Han YP, Song YJ, et al. DNA microarray analysis of genome dynamics inYersiniapestis: insight into bacterial genome microevolution and niche adaptation[J]. Med J CPLA, 2004,29(3): 204-210. (in Chinese)
周冬生, 韩延平, 宋亚军,等. 鼠疫耶尔森氏菌基因组进化与生态位适应研究[J]. 解放军医学杂志,2004,29(3): 204-210.
[5]Wang ZY, Luo SD, Yu XT, et al. The discovery and research ofMicrotusfuscusplague natural foci in Qinghai province[J]. China J Endem, 2004, 23(1): 69-72. (in Chinese)
王祖郧,罗松达卫,于晓涛,等.青海省青海田鼠鼠疫自然疫源地的发现与研究[J]. 中国地方病防治杂志, 2004, 23(1): 69-72.
猜你喜欢
作文大王·低年级(2021年11期)2021-12-22 00:58:13
中国药学药品知识仓库(2021年12期)2021-12-07 00:43:43
小天使·一年级语数英综合(2021年2期)2021-05-08 07:48:34
小学生(看图说画)(2020年4期)2020-04-01 11:58:18
江苏农业科学(2019年3期)2019-08-10 04:20:00
中国学术期刊文摘(2019年23期)2019-01-29 11:49:47
农家科技中旬版(2018年9期)2018-10-14 13:09:36
小学教学研究(2016年13期)2016-04-16 05:17:25
中国人兽共患病学报(2015年12期)2015-01-25 03:21:47
小天使·一年级语数英综合(2014年2期)2014-01-07 00:34:22
