
多房棘球绦虫EM95蛋白的生物信息学分析
2014-04-09 03:18范彦雷娄忠子闫鸿斌贾万忠
中国人兽共患病学报 2014年4期
范彦雷,李 立,娄忠子,闫鸿斌,贾万忠
泡型棘球蚴病(alveolar echinococcosis, AE),又称泡型包虫病,是由多房棘球绦虫(Echinococcusmultilocularis, Em)的幼虫引起的一种危害严重的人畜共患寄生虫病,可感染包括人在内的多种动物,呈世界性分布。目前该病呈现感染地区以及感染宿主扩增的趋势,在一些非流行区的国家和地区也首次出现有感染人的报道[1],同时也有该寄生虫感染非特异性宿主的报道,从而间接增加人的感染[2]。在我国的青海、新疆、甘肃、宁夏及四川等地区,人泡型棘球蚴病又称“虫癌”,严重威胁着广大牧民的生命和健康[3]。然而目前对于泡型棘球蚴病的治疗以手术治疗为主,药物治疗为辅,但在术后易出现疾病的复发现象,效果欠佳。其防治主要是给犬定期驱虫和宣传教育为主的综合防治措施,在偏远地区该防治措施的效果也不甚理想。因此当务之急应积极寻找新的有效防治措施来预防和控制该病的流行和进一步蔓延。
免疫预防是防止各种疾病流行方法中比较有效的措施,泡型棘球蚴病也不例外。目前,多房棘球绦虫相关抗原分子主要有EMⅡ/3、EM10、EM14-3-3、EM18、EMY162,然而当前研究结果表明,这些抗原分子的保护力较低或不具备其坚强保护力[4-7]。因此需要进一步研究新型的多房棘球绦虫抗原分子作为候选疫苗分子。EG95分子是一种分子量为24.5 ku的天然抗原[8],在细粒棘球绦虫的各个阶段均有表达,是针对各阶段均具有有效保护作用的疫苗分子[9]。EG95重组抗原、合成肽疫苗、DNA疫苗、重组酵母、重组牛痘病毒疫苗、重组BCG疫苗等的实验结果均表明EG95抗原对细粒棘球绦虫感染宿主具有很好的保护效果,是目前最有效的抗细粒棘球绦虫的疫苗分子之一[10]。EM95基因最早由Gauci 克隆和鉴定的,其全长2 34 bp,含有3个外显子和2个内含子,其结构与EG95基因相似[11-12]。对其核苷酸和氨基酸序列的进一步分析表明,EM95是一种胞外型和分泌型蛋白,是成为疫苗分子的先决有效条件[11]。EM95免疫小鼠后期保护性可达到78.5%~82.9%,但低于EG95对绵羊的保护率[11,13-14]。因此,本文利用生物信息软件从生物信息学方面对EM95B细胞抗原表位进行分析,并与EG95蛋白分子的一级结构进行比对,为制备更为有效的EM95疫苗的相关实验研究奠定理论基础和提供指导。
1 材料与方法
1.1实验材料 从美国国家生物技术信息中心(National Center for Biotechnology, NCBI)数据库中检索EM95蛋白序列,确定要分析的EM95蛋白序列(登录号:AAL51153)和EG95蛋白序列(登录号:AAQ93497)。
1.2生物信息学数据库和软件 利用常见生物信息学分析网站和本地分析软件(见表1)对EM95蛋白的二级结构、跨膜结构域以及B细胞抗原表位等进行综合分析与预测,同时比对了EM95和EG95两个蛋白一级序列并分析了差异位点存在的区域。
2 结 果
EM95蛋白质序列由Gauci等提交,其序列来源于从德国普通小鼠中发现的原头节所提取的mRNA所翻译的蛋白质。由156个氨基酸残基组成,属分泌蛋白,含有一个糖基磷脂酰肌醇(GPI)锚定点和一个Fn-3型结构域及N∕C端疏水区[15]。此外有此结构特征的蛋白质还有EG95、To45W、TSOL16、TSOL18等[16]。其中Fn-3型结构域是六钩蚴与中间宿主免疫内分泌间相互作用的重要区域,在诱导免疫保护方面起到重要作用。
2.1EM95和EG95氨基酸残基比对及差异位点分析 利用EBI网站中的ClustalW2软件对EM95和EG95氨基酸残基进行比对(见图1),并统计差异位点的氨基酸残基(见表2),统计数据显示,EM95和EG95蛋白156个氨基酸残基中存在31个突变位点,差异达22.2%,其中极性氨基酸残基突变为非极性的有3个位点,非极性氨基酸残基突变为极性的有4个位点,其余均为极性与极性、非极性与非极性间突变。
表1 常用生物信息学网站及软件

图1利用ClustalW2软件对EM95和EG95蛋白氨基酸残基比对
Fig.1AminoacidresiduealignmentofEM95andEG95proteinusingClustalW2

表2 EM95和EG95蛋白氨基酸残基差异位点分析
注:数字表示序列比对差异位点;+∕-分别表示氨基酸残基极性内与极性间的变异
Note: Numbers indicated the alignment different position; +∕- represents the amino acid residue divergence of intra- and inter-polarity.
2.2EM95抗原B表位分析
2.2.1EM95抗原蛋白质二级结构预测 利用EXPASY服务器中SOPMA在线软件对EM95二级结构进行预测,EM95蛋白质中柔性结构占氨基酸残基总数的45.51%,其中无规卷曲占39.10%,β转角占6.41%;α螺旋和β折叠(extended strand)分别占25.64%和28.85%,各种结构在EM95抗原中分布情况(见图2)。
2.2.2EM95蛋白跨膜区预测 使用TMHMM对EM95蛋白进行跨膜区预测,TMHMM综合了跨膜区疏水性、电荷偏倚、螺旋长度和膜蛋白拓扑学限制等性质,采用隐马氏模型,对跨膜区及膜内外区进行整体的预测(见图3)。从预测结果可以看出EM95蛋白存在一个跨膜区,其中1~132位氨基酸残基位于细胞外,139~155位氨基酸残基跨膜,156位氨基酸残基位于细胞内。这与EM95蛋白C端是疏水区相吻合。
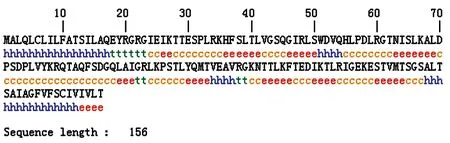
图2SOPMA分析的EM95二级结构
Fig.2ThesecondarystructureofEM95usingSOPMAanalysis

图3 EM95蛋白质跨膜结构区预测
Fig.3ThetransmembranestructurepredictionofEM95protein
2.2.3DNAStar软件分析EM95蛋白 DNAStar软件对EM95蛋白的预测(见图4),Kyte-Doolittle方法预测EM95亲水性区域为:15~37、47~64、69~87、94~101和105~134。Eisenberg方法预测的两亲性结果在亲水区域范围内,与Kyte-Doolittle方法相吻合。Karplus-Schulz预测EM95柔性区域显示可塑性较强的区域为:18~35,41~47,57~64,69~75,78~82,85~88,93~97,108~122,125~138;采用Jameson-Wolf方法预测EM95蛋白抗原性发现存在5个高度的抗原区,分别位于17~35,57~64,67~89,92~98,105~131。表面区域通常是良好的抗原,在抗体产生上会有更好的效果,Emni方法表明,EM95蛋白在17~20,27~34,76~82,109~114,126~131等部位氨基酸残基存在突出表面区。综合以上数据可推测出EM95蛋白存在5个B细胞抗原表位,分别为:17~35,47~64,69~89,92~98,105~131。
2.2.4在线网站IEBD预测EM95蛋白抗原B细胞表位 利用在线网站IEBD预测EM95蛋白抗原细胞表位(见图5):1)Chou﹠Fasman Beta-Turn Prediction分值较高区域为(18~24、28~35、41~49、51~65、68~77、83~90、93~100、108~113、127~129、132~139);2)对EM95进行Emini表面可及性分析,(分值较高区域:17~20、27~36、69~72、75~83、94~95、107~121、126~130);3)Karplus﹠Schulzz柔韧性区域预测,EM95分值高的区域为:18~35、42~47、57~64、69~75、78~82、85~88、93~98、108~122、125~139;4)Kolaskar ﹠ Tongaonkar 抗原性指数预测,可能存在的表位区域为:4~18、33~48、50~59、65~79、86~94、96~107、137~152;5)Parker的亲水性预测显示EM95的高亲水性区域为:18~33、45~47、59~65、68~73、75~76、79~89、103~116、118~121、124~139;6)Bepipred抗原表位预测,EM95存在多个Bepipred线性抗原表位区域为:19~21、25~33、59、61、69~77、81~86、107~111、127~134、137~138。综合以上参数分析,得出EM95蛋白可能存在7个潜在B细胞抗原表位区域:18~35,41~48,50~65,69~90,93~100,107~121,124~139。

图4 DNAStar软件分析EM95蛋白各项参数
综合以上两个B细胞抗原表位预测分析软件结果,最终确定EM95抗原B细胞表位有6个可能区域,分别为:17~35、42~64、69~90、92~100、105~121、124~139。
3 讨 论
随着生物信息学的快速发展,计算机辅助疫苗设计技术正带动着生物技术产业向一个新领域飞速发展。目前在线或本地蛋白质分析工具为科研工作者分析蛋白质序列,寻找蛋白质序列中的特殊结构,进而对其进行加工和修饰,从而使其获得免疫原性更强的抗原。自上世纪80年代Hoop和Woods提出亲水性参数预测抗原表位方法以来[17],已有许多参数、算法发表,在B细胞蛋白抗原表位预测研究中起到了巨大的推动作用。Hoop-Woods方案认为蛋白质氨基酸残基可分为亲水性和疏水性残基两类,该方案是以氨基酸残基由有机相到水相环境转移的自由能为依据来计算氨基酸的亲水性,亲水性残基一般位于蛋白质表面,而疏水性残基则位于蛋白质内部,位于蛋白质表面的亲水性氨基酸残基易于B细胞膜上的的受体(BCR)结合,进而刺激B细胞活化产生抗体。Emini表面可及性分析方案放映了抗原内、外两层氨基酸残基的分布情况[18],最早在分析甲型肝炎病毒(HAV)和脊髓灰质炎Ⅰ型病毒VP1蛋白表面特征时,合成一段含有HAV特异性氨基酸序列能够诱导产生抗HAV中和抗体的合成肽,发现这两个病毒蛋白质间存在结构同源特征[19]。Karplus和Schulzz柔韧性区域预测是基于已知结构的31个蛋白质的Cx的温度β因子,认为蛋白质抗原多肽链骨架具有一定的活动性,其活动性强的区域易成为抗原表位来预测蛋白质抗原表位的方法[20]。Kolaskar和Tongaonkar 抗原性指数预测方案依据氨基酸残基理化特征及在已知表位片段上出现的频率而发展起来的预测蛋白质抗原决定簇的方法,其预测率达75%[21];Jameson-Wolf抗原指数预测方案则是通过对20个研究深入的蛋白质中的69个连续位点上606个氨基酸残基统计分析而建立起来抗原性刻度[22]。Parker亲水性指数预测依据20个合成肽模型保留时间的高效液相层析参数,同时比较了其他9个相同尺度范围的参数,结果表明,该参数与抗原性吻合度最好,同时结合其他三个预测抗原性较好的参数来提高预测的准确度[23]。Bepipred线性抗原表位预测方案是在最好的预测线性B细胞表位的隐马尔科夫模型的基础上发展起来的抗原表位预测方法,其预测效果较其他方法都佳[24]。此外,二级结构预测方案认为β转角多为位于蛋白质抗原表面的凸出结构,易与抗体发生特异性结合,而α螺旋、β折叠结构则不易与抗体嵌合,因此β转角区域可能为抗原表位[25]。由于各个分析软件依据的参数及模型不一样,因此只有综合各个预测结果才能提高抗原表位预测的准确度。
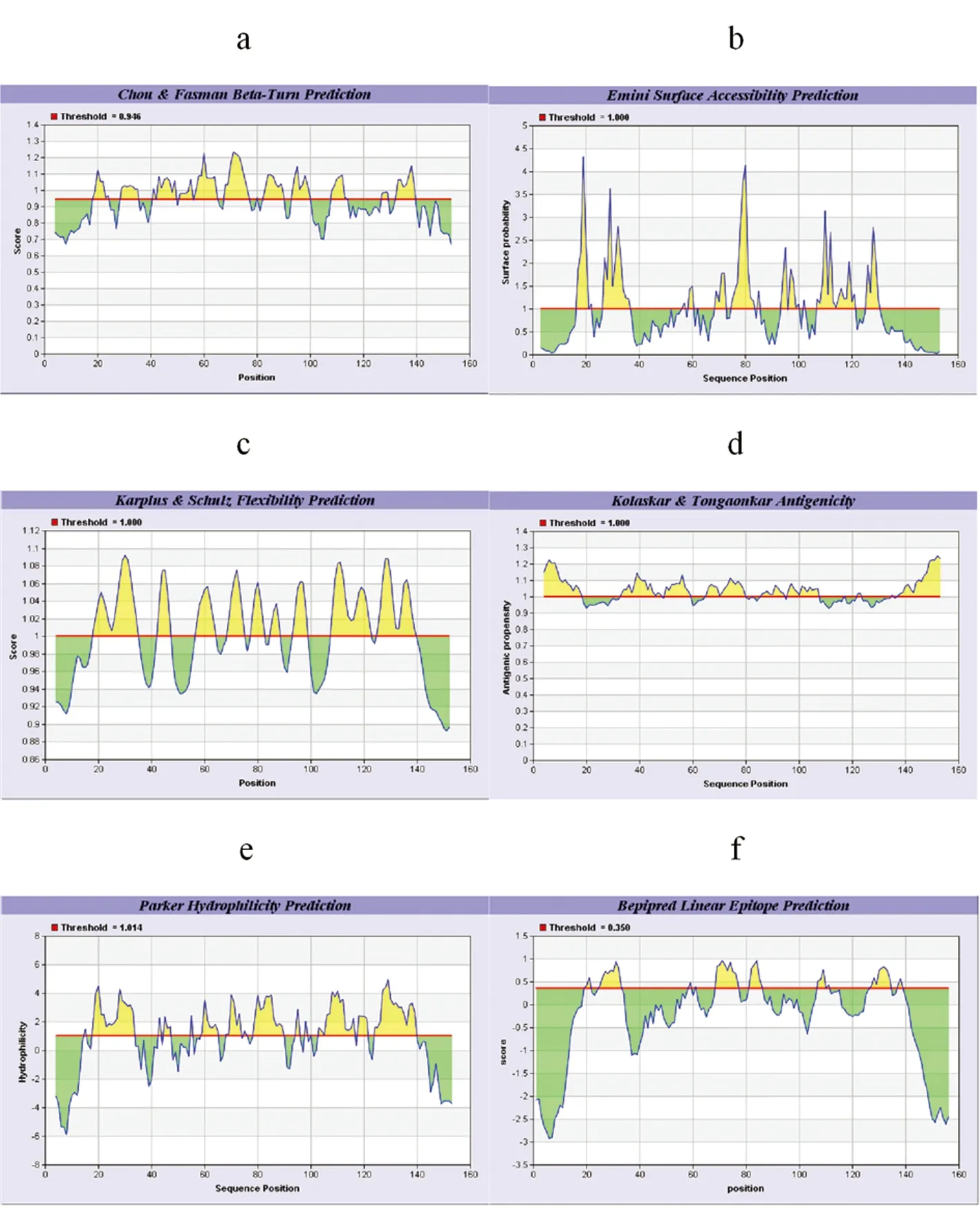
图5IEDB分析EM95的B细胞抗原表位
a: Chou ﹠Fasman β转角预测;b: Emini 表面可及性预测;c: Karplus ﹠Schulzz柔韧性区域预测;d: Kolaskar ﹠ Tongaonkar 抗原性指数预测;e: Parker的亲水性指数预测;f:Bepipred线性抗原表位预测
Fig.5TheparametersofEM95usingIEDBanalysis
a: Chou ﹠ Fasman Beta-Turn Prediction;b: Emini Surface Accessibility Prediction; c: Karplus ﹠ Schulzz Flexibility Prediction; d: Kolaskar ﹠ Tongaonkar Antigenicity Prediction; e: Parker Hydrophilicity Prediction; f:Bepipred Linear Epitope
EG95蛋白抗原作为免疫疫苗在保护中间宿主中获得了很好的效果,EG95克隆重组抗原免疫中间宿主(羊),在六钩蚴感染实验中获得96%~98%免疫保护效果[14];Larrieu等[26]利用EG95蛋白抗原免疫了3 146只羔羊和311条家犬,同样获得了理想的保护效果,并进一步证明EG95蛋白抗原疫苗能保护至3岁龄的动物;同样EG95蛋白抗原在免疫牛时也取得了理想的免疫效果[27]。而EM95蛋白抗原免疫中间宿主(啮齿动物)时,获得的保护率低于EG95蛋白抗原免疫羊所取得的保护率。因此对EM95蛋白和EG95蛋白二级结构及抗原表位分析将有助于找出两种抗原的差异,为设计高效的EM95蛋白抗原提供思路。李玉娇等[8]分析了EG95蛋白抗原的二级结构及抗原表位,最终预测EG95蛋白可能存在7个潜在B细胞抗原表位,本研究利用了同样的分析工具,对EM95蛋白进行抗原表位预测,结果显示EM95蛋白可能存在6个抗原表位区域。同时比对了两种蛋白质的氨基酸残基序列,发现在预测EG95蛋白抗原表位7个区域中与EM95蛋白相对应的位置均存在不同程度的氨基酸残基替换,其中第1个抗原表位区域存在2个氨基酸残基极性间替换(M替换为R、T替换为I),第3个潜在表位存在1个(S替换为P),第4个潜在抗原表位存在1个(T替换为A),第6个潜在抗原表位存在1个(I替换为T)。氨基酸残基极性间的替换可能对蛋白质的空间结构造成很大影响,分析EM95蛋白和EG95蛋白潜在抗原表位区域氨基酸的残基差异将有助于阐释EM95蛋白抗原保护力低于EG95蛋白的原因,为开发理想的EM95蛋白抗原疫苗奠定理论依据和提供指导。
4 小 结
本研究利用生物信息学工具对EM95蛋白的二级结构、跨膜区以及B细胞抗原表位进行了综合预测,同时分析了EM95和EG95蛋白间的差异,为研发具有高效保护率的EM95蛋白抗原疫苗提供理论基础及预防泡型包虫病的流行奠定实验基础。
参考文献:
[1]van Dommelen L, Stoot JH, Cappendijk VC, et al. The first locally acquired human infection ofEchinococcusmultilocularisin the Netherlands[J]. J Clin Microbiol, 2012, 50(5): 1818-1820. DOI: 10.1128/JCM.06355-11
[2]Karamon J, Sroka J, Cencek T. The first detection ofEchinococcusmultilocularisin slaughtered pigs in Poland[J]. Vet Parasitol, 2012, 185(2-4): 327-329. DOI: 10.1016/j.vetpar.2011.09.022
[3]Eckert J, Gemmell MA, Meslin FX, et al. WHO/OIE Manual on echinococcosis in humans and animal: a public health problem of global concern[M]. Paris: WHO/OIE, 2001: 47-58.
[4]Yang M, Li WG. Changes of cytokines in mice immunized with recombinant Bb-EmII/3-Em14-3-3 vaccine ofEchinococcusmultilocularis[J]. Chin J Cell Mol Immunol, 2008, 24(8): 781-784. (in Chinese)
杨梅, 李文桂. 多房棘球绦虫重组Bb-EmII/3-Em14-3-3疫苗诱导小鼠细胞因子变化的研究[J]. 细胞与分子免疫学杂志, 2008, 24(8): 781-784.
[5]Yang M, Li WG, Zhu YM. Construction and expression efficiency of recombinant Bb-Em14-3-3 vaccine ofEchinococcusmultilocularis[J]. J Xi’an Jiaotong Univ (Med Sci), 2008, 29(2): 148-151. (in Chinese)
杨梅, 李文桂, 朱佑明. 多房棘球绦虫重组Bb-Em14-3-3疫苗的构建及其表达效率[J].西安交通大学学报, 2008, 29(2): 148-151.
[6]Li WG, Chen YT. Research progress on the Em18 ofEchinococcusmultilocularis[J]. Chin J Zoonoses, 2009, 25(1): 67-69. (in Chinese)
李文桂, 陈雅棠. 多房棘球绦虫Em18研究进展[J]. 中国人兽共患病学报, 2009, 25(1): 67-69.
[7]Kouguchi H, Matsumoto J, Katoh Y, et al. The vaccination potential of EMY162 antigen againstEchinococcusmultilocularisinfection[J]. BBRC, 2007, 363(4): 915-920.DOI: 10.1016/j.bbrc.2007.09.023
[8]Li YJ, Wang J, Zhao H, et al. Bioinformatics prediction on Eg95 antigen epitopes ofEchinococcusmultilocularis[J]. Chin J Zoonoses, 2011, 27(10): 892-896. (in Chinese)
李玉娇, 王晶, 赵慧, 等. 细粒棘球绦虫Eg95抗原表位的生物信息学预测[J]. 中国人兽共患病学报, 2011, 27(10): 892-896.
[9]Zhang W, Li J, You H, et al. Short report:Echinococcusgranulosusfrom Xinjiang, PR China: cDNAS encoding the EG95 vaccine antigen are expressed in different life cycle stages and are conserved in the oncosphere[J]. Am J Trop Med Hyg, 2003, 68(1): 40-43.
[10]Li WG, Chen YT. Research progress on the Eg95 vaccine ofEchinococcusgranulosus[J]. Chin J Zoonoses, 2006, 22(11): 1070-1072. (in Chinese)
李文桂, 陈雅棠. 细粒棘球绦虫Eg95疫苗研究进展[J]. 中国人兽共患病学报, 2006,22(11): 1070-1072.
[11]Gauci C, Merli M, Muller V, et al. Molecular cloning of a vaccine antigen against infection with the larval stage ofEchinococcusmultilocularis[J]. Infect Immun, 2002,70(70): 3969-3972. DOI: 10.1128/IAI.70.7.3969-3972.2002
[12]Chow C, Gauci CG, Cowman AF, et al. A gene family expressing a host-protective antigen ofEchinococcusgranulosus[J]. Mol Biochem Parasitol, 2001, 118(1): 83-88.DOI: 10.1016/S0166-6851(01)00373-5
[13]Lightowlers MW, Jensen O, Fernandez E, et al. Vaccination trials in Australia and Argentina confirm the effectiveness of the EG95 hydatid vaccine in sheep[J]. Int J Parasitol, 1999, 29(4): 531-534. DOI: 10.1016/S0020-7519(99)0000-X
[14]Lightowlers MW, Lawrence SB, Gauci CG, et al. Vaccination against hydatidosis using a defined recombinant antigen[J]. Parasite Immunol, 1996, 18(9): 457-462. DOI: 10.1111/j.1365-3024.1996.tb01029.x
[15]Kyngdon CT, Gauci CG, Gonzalez AE, et al. Antibody responses and epitope specificities to theTaeniasoliumcysticercosis vaccines TSOL18 and TSOL45-1A[J]. Parasite Immunol, 2006, 28(5): 191-199. DOI: 10.1111/j.1365-3024.2006.00820.x
[16]Waterkeyn J, Gauci C, Cowman A, et al. Sequence analysis of a gene family encodingTaeniaovisvaccine antigens expressed during embryogenesis of egg[J]. Mol Biochem Parasitol, 1997, 86(1): 75-84. DOI: 10.1016/S0166-6851(97)02851-X
[17]Hoop JP, Wood KR. Prediction of protein antigenic determinants from amino acid sequences[J]. Immunology, 1981, 78(6): 3824-3828.
[18]Florea L, Halldòrsson B, Kohlbacher O, et al. Epitope prediction algorithms for peptide-based vaccine design[J]. Proc IEEE Comput Soc Bioinform Conf, 2003, 2: 17-26. DOI: 10.1109/CSB.2003.1227293
[19]Emini EA, Hughes JV, Perlow DS, et al. Induction of hepatitis A virus-neutralizing antibody by a virus-specific synthetic peptide[J]. J Virol, 1985, 55(3): 836-839.
[20]Karplus PA, Schulz GE. Prediction of chain flexibility in proteins[J]. Naturwissenschaften, 1985, 72(4): 212-213. DOI: 10.1007/BF01195768
[21]Kolaskar AS, Tongaonkar PC. A semi-empirical method for prediction of antigenic determinants on protein antigens[J]. FEBS Lett, 1990, 276(1-2): 172-174. DOI: 10.1016/0014-5793(90)80535-Q
[22]Le SI, Thedja MD, Roni M, et al. Prediction of conformational changes by single mutation in the hepatitis B virus surface antigen (HBsAg) identified in HBsAg-negative blood donors[J]. Virol J, 2010, 18(7): 326-334. DOI: 10.1186/1743-422X-7-326
[23]Parker JM, Guo D, Hodges RS. New hydrophilicity scale derived from high-performance liquid chromatography peptide retention data: correlation of predicted surface residues with antigenicity and X-ray-derived accessible sites[J]. Biochemistry, 1986, 25(19): 5425-5432. DOI: 10.1021/bi00367a013
[24]Larsen JE, Lund Q, Nielsen M. Improved method for predicting linear B-cell epitopes[J]. Immunome Res, 2006, 2: 2. DOI: 10.1186/1745-7580-2-2
[25]Lai LH, Xu XJ, Tang YQ. The structure prediction and molecular designing of protein[J]. Univ Chem, 1993: 49. (in Chinese)
来鲁华, 徐筱杰, 唐有祺. 蛋白质的结构预测与分子设计[J]. 大学化学, 1993: 49.
[26]Larrieu E, Herrero E, Mujica G, et al. Pilot field trial of the EG95 vaccine against ovine cystic echinococcosis in Rio Negro, Argentina: Early impact and preliminary data[J]. Acta Trop, 2013, 127(2): 143-151. DOI: 10.1016/j.actatropica.2013.04.009
[27]Heath DD, Robinson C, Lightowlers MW. Maternal antibody parameters of cattle and calves receiving EG95 vaccine to protect againstEchinococcusgranulosus[J]. Vaccine, 2012, 30(50): 7321-7326. DOI: 10.1016/j.vaccine.2012.08.076
猜你喜欢
生物化学与生物物理进展(2022年7期)2022-07-25
生物化学与生物物理进展(2022年6期)2022-07-21
食品与生物技术学报(2021年5期)2021-06-24
中学生数理化(高中版.高考理化)(2021年2期)2021-03-19
智慧健康(2021年2期)2021-03-15
昆明医科大学学报(2021年1期)2021-02-07
温州医科大学学报(2019年4期)2019-04-28
中国免疫学杂志(2017年1期)2017-01-17
肿瘤影像学(2015年3期)2015-12-09
癌变·畸变·突变(2015年3期)2015-02-27
