
辽东山地山杨天然次生林空间结构分析
2014-03-26 07:49陈丽华郭文体李胜男高洪翠李长暄李海燕
西北农林科技大学学报(自然科学版) 2014年8期
周 娟,陈丽华,郭文体,李胜男,高洪翠,李长暄,李海燕
(1 北京林业大学 水土保持学院,北京 100083;2 中国科学院 沈阳应用生态研究所,辽宁 沈阳 110016;3 辽宁老秃顶子国家级自然保护区抚顺管理局,辽宁 抚顺 113208)
林分空间结构是指林木在林地上的分布格局及其属性在空间上的排列方式,也就是林木之间树种、大小、分布等空间关系,林分空间结构在很大程度上决定了林分的稳定性、发展的可能性和经营空间的大小[1-3]。林分的空间结构反映了森林群落内物种间的空间关系,即林分中林木的水平分布格局和空间排列方式,其有助于对混交林的经营措施进行合理评价,有助于新的林分生长模型研究,同时还有助于森林经营方案的制定[4]。因此,近年来对林分空间结构的研究倍受关注,提出了多种描述林分空间结构的参数[5-6],如树种空间隔离程度(即混交)、林木个体在水平面上的分布形式(即林木空间分布格局),分别采用混交度和角尺度进行表征[7-8]。
本研究通过胸径结构分析、树高结构分析、混交度和角尺度等,对辽宁省老秃顶子自然保护区内的天然山杨林进行了空间结构分析,以了解林分的结构,为使其发挥最大功能提供依据,为制定和实施森林经营及保护规划提供可释性强的空间结构信息支持。
1 研究区概况
研究区位于辽宁省东部的抚顺市老秃顶子国家级自然保护区,地理坐标为124°41′13″~125°05′15″E,41°11′11″~41°21′34″ N,总面积15 217.3 hm2。区内土壤以棕壤和暗棕壤为主,pH值5.15~6.12,棕壤为本区的地带性土壤,主要分布于海拔900 m以下的落叶阔叶林中,暗棕壤为本区的垂直地带性土壤,主要分布于海拔900 m以上的中山草甸和暗针叶林中,土壤湿润,有机质含量高;气候属北温带大陆性季风气候,由于受海洋气候的影响,雨量充沛,年降水量900~1 200 mm,且多集中在6-8月份,相对湿度67%,极端最高气温37.2 ℃,极端最低气温-37.5 ℃,年平均气温5.2 ℃,年无霜期133 d。保护区内水资源丰富,共有大小河流17条,其中老秃顶子山西麓的3条河流属太子河水系,是辽宁省第三大河太子河的发源地,本区还是辽河的发源地和鸭绿江的重要源头之一,为重要的水源涵养林区。区内植物群落组成复杂,森林覆盖率达到97%,并保存了较为完整的植物垂直分布带谱。森林以天然次生林和人工林为主,主要乔木树种有蒙古栎(Quercusmongolica)、白桦(Betulaplatyphylla)、油松(Pinustabulaeformis)、落叶松(LarixgmeliniiRupr.)、山杨(Populusdavidiana)、花楷槭(AcerukurunduenseTrautv. et Mey.)、胡桃楸(JuglansmandshuricaMaxim.)、色木槭(AcermonoMaxim)、千金榆(Carpinuscordata)等。
2 研究方法
2.1 野外调查
本研究试验地设置在辽宁省抚顺市老秃顶子国家自然保护区内,于2012年7-8月在保护区内选择具有代表性的以山杨为优势种的天然次生林作为研究对象,采用固定样地调查法进行调查。选择3块标准地(50 m×50 m),采用相邻网格法,将样地划分为10 m×10 m的小样方,调查时将每个小样方的西南角设定为坐标原点,用皮尺分别测量调查单元中每株树木的位置坐标(x,y),x、y分别表示东西方向和南北方向的坐标。用GPS对样地边界上每个网格结点进行定位,测定样地坡度、坡向、经纬度、海拔、地形地貌等立地因子,并记录树木的名称、坐标、树高、胸径、冠幅、枝下高、优势度、损伤状况、干形质量、病虫害和起源等。
2.2 数据分析
按照森林资源清查和森林经理的观点,从3个方面入手完整描述天然林的空间结构:①树种的空间隔离程度(非同质性);②林木个体大小分化程度(非均一性);③林木个体空间分布格局(非规则性)[9]。空间结构分析以空间结构单元分析为基础。空间结构单元由任意1 株单木与相邻木和参照树构成的结构关系有5种,即零度、弱度、中度、强度和极强度[10]。本研究采用胸径和树高结构特点描述林木在水平或垂直方向上的竞争状态,用树种混交度描述空间隔离程度,用角尺度描述林分水平分布格局。
(1)胸径和树高结构分析。《森林资源规划设计调查主要技术规定》中提到,根据林分平均直径确定径阶距,一般当林分平均直径在12 cm以上时,以4 cm 为一个径阶;6~12 cm 时,以2 cm 为一个径阶;小于6 cm 时,可以采用1 cm 为1个径阶[11]。本研究中的4个树种(山杨、蒙古栎、黄桦、刺楸)平均胸径都在12 cm以上,因此胸径径阶距采用4 cm。根据实际调查,树高距采用2 m。
(2)树种混交度(Mi)。指参照树i的4株最近相邻木中与参照树不属于同种的个体所占的比例。计算公式为[10]:
式中:n为相邻木的总株数,n=4;当参照树i与相邻木j为同种时,Vij=0;为非同种时,Vij=1。因此,Mi有5种可能取值,即0,0.25,0.5,0.75和1,其分别对应混交度的描述为零度、弱度、中度、强度和极强度混交。

式中:n为相邻木的总株数,n=4;当α角小于标准角α0,则Zij=1,否则Zij=0。
式中,N指参照树的总株数。为了能明显地表达出混交度等结构参数的林学意义,本研究选用n=4,即在参照树周围选择4株相邻木组成一个结构框架。研究表明,由1株参照树与4株相邻木所组成的结构单元中,参照树与4株最近相邻木构成的结构关系有5种:零度、弱度、中度、强度和极强度;n= 3或n= 5时,在类型划分上仅有4种或6种,缺乏中间过渡类型,不符合自然规律;n=4时,过渡阶段更加完整,生物学意义更加明显,空间结构信息较完整,且这种结构单元的可释性和可操作性都较强,适宜于描述林分的空间结构特征[14-15]。
3 结果与分析
3.1 山杨天然次生林的树种组成
山杨天然次生林物种丰富度较高,本研究只选取了在数量上有一定优势的树种。由表1可以看出,山杨天然次生林样地内乔木层主要有种群11个,3个样地的林分密度分别为683,849和774 株/hm2,其中就数量、胸径、树高综合来看,山杨均为该林分的优势树种,其主要伴生树种有蒙古栎、花楷槭、色木槭、黄桦、刺楸、千金榆等,除这些树种外,其他伴生树种如核桃楸、糠椴、怀槐等株数比例都非常小,因此没有纳入本研究中。样地1中,大叶杨的胸径和树高都是最大的,但数量相对较少,只有66株/hm2;刺楸在生长上不占优势,平均树高和平均胸径都是最小的。在样地2中,山杨是绝对的优势树种,而花楷槭和千金榆则在竞争中处于不利地位,胸径和树高都普遍偏小;在样地3中,刺楸相对是弱势群体,数量少,且胸径、树高都是最小的。

表 1 老秃顶子国家自然保护区山杨天然次生林样地中各树种林分因子的统计

续表 1 Continued Table 1
3.2 山杨天然次生林的胸径和树高结构
3.2.1 胸径结构特点 不同种类的树木胸径分布有不同的特点,本研究选取数量较多的4种林木,分别是山杨、蒙古栎、黄桦和刺楸,径阶距采用4 cm,其胸径分布特点见图1。
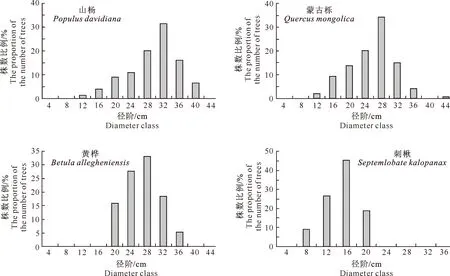
图 1 老秃顶子国家自然保护区山杨天然次生林中4种主要林木胸径的径阶分布
由图1可以看出,这4个树种的胸径分布特征表现出一致性,整体上都呈现正态分布,只不过山杨和蒙古栎的胸径结构相对更加复杂,从12~40 cm径阶都有分布,而黄桦集中在较大径阶(20~36 cm),与之相反,刺楸多分布在较小径阶(8~20 cm)。分别来看每个树种,山杨在32 cm径阶的株数最多,占到总株数的31.56%,其次是28 cm径阶,比例为20.00%;蒙古栎株数最多的是在28 cm这个径阶,比例达到34.06%,而在4 cm和8 cm这2个径阶上没有树木分布,其最大径阶达到44 cm;黄桦的胸径主要集中在24 cm和28 cm这2个径阶上,比例之和超过了50%;刺楸的胸径分布相对来说比较简单,只在4个径阶有分布,且以16 cm径阶的株数最多。不难看出,作为优势树种的山杨在水平方向上竞争优势最大。
3.2.2 树高结构特点 树高距采用2 m,4个主要树种的树高分布特点见图2。图2显示,4个树种的树高结构及其分布特点与胸径结构有相似之处,总的趋势是随着林分树高阶的增大,其所占株数的比例先增大后减小,表现为正态分布的特点。其中山杨和黄桦以22 m树高阶的株数最多,蒙古栎以20 m树高阶的株数最多,而刺楸株数最多的树高阶在12 m处。这说明在垂直方向上,山杨仍处于竞争优势状态,而刺楸在垂直生长上处于相对劣势,其在乔木层的较下层,能够接受的阳光有限,因而生长也会受到一些影响。结合胸径和树高分布可以看出,4个树种之间存在密切的联系,胸径大的树木往往长得也比较高。
3.3 山杨天然次生林林木的种间结构
混交度反映了林分的种间隔离程度,一般认为混交度越大,林分相对越稳定。从图3可以看出,样地1、样地2、样地3分别以强度混交、中度混交和极强度混交的比例为最大。综合来看,3块样地的平均混交度都比较高,这说明同种树种聚集在一起的情况相对比较少,多数树种不与本树种为伴,林分比较稳定。
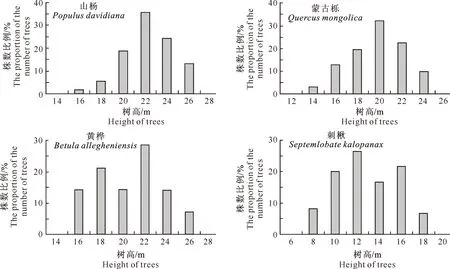
图 2 老秃顶子国家自然保护区山杨天然次生林中4种主要林木树高的径阶分布
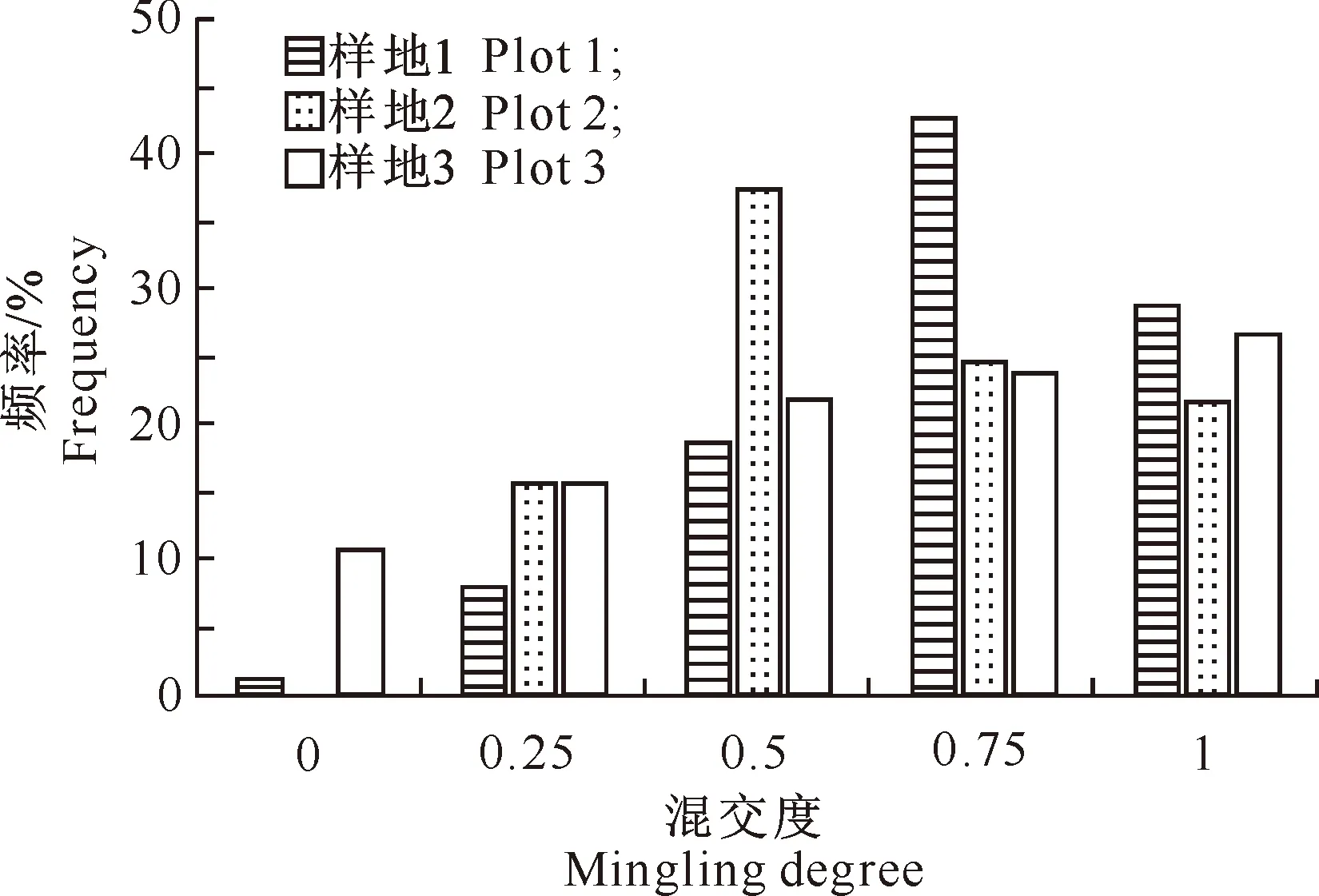
图 3 老秃顶子国家自然保护区山杨天然次生林中各样地林分的混交度分布
为了得出各树种的混交度信息,进一步对样地数据分析得出表2。由表2可知,各树种平均混交度依次是:紫椴(0.90)>黄桦(0.89)>大叶杨(0.86)>色木槭(0.82)>蒙古栎(0.79)>花曲柳(0.75)>白桦(0.71)>花楷槭(0.69)>刺楸(0.67)>千金榆(0.55)>山杨(0.45)。山杨为此天然次生林中的优势树种,其平均混交度为0.45,说明山杨周围常有2株相同树种伴生,与其他树种比较其混交度是最小的。研究还发现,千金榆、刺楸和花楷槭这3个树种的混交度也比较小,究其原因可能是由于这3个树种的树高、胸径都比较小,有的就两三株簇生在一起,导致周围同种树较多,混交度小。黄桦、大叶杨、色木槭和紫椴的平均混交度都超过了0.8,说明树种很少呈现单种聚集,其周围至少有3株其他树种伴生,这与调查情况相符,其各自株数不是很多,基本都零星均匀分布在整个样地中。
除山杨外,其余树种呈零度混交和弱度混交的株数频率之和均没有超过20%,且全林分平均混交度达到0.65,这说明天然山杨林混交程度较高,经营保护较好,树种组成结构多样,林分比较稳定。

表 2 老秃顶子国家自然保护区山杨天然次生林中各树种的混交度及其频率分布
3.4 山杨天然次生林林木的空间分配格局
用角尺度描述林分中的林木个体分布格局时,关注的是林木个体之间的方位关系。根据角尺度的定义,Wi值越大表示参照树周围的相邻木分布越不均匀。由表3可知,所有树种基本以随机分布的比例最大,而绝对均匀分布和绝对不均匀分布所占比例都比较小。
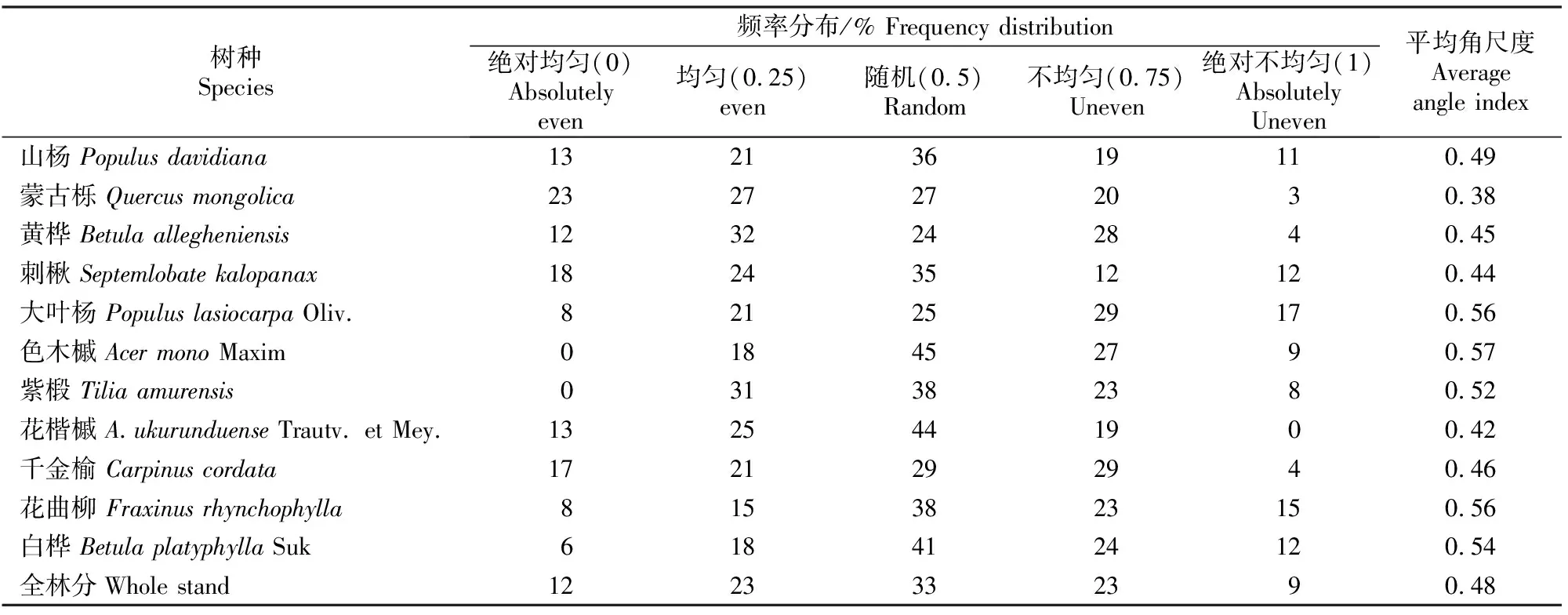
表 3 老秃顶子国家自然保护区山杨天然次生林中各树种的角尺度及其频率分布
对于4株最近相邻木而言,随机分布的角尺度取值范围是[0.475,0.517],因此平均角尺度小于0.475 的分布就是均匀分布,大于0.517 就是团状分布,根据这个原则来判断林木个体的空间分布格局。其中蒙古栎、黄桦、刺楸、花楷槭和千金榆的平均角尺度均小于0.475,说明其整体呈均匀分布;再看整个林分的优势树种山杨,其平均角尺度为0.49,介于 0.475 与0.517,是随机分布,且绝对均匀和绝对不均匀分布的比例之和只占24%;大叶杨、色木槭、紫椴、花曲柳和白桦的平均角尺度均比0.517要稍大,说明其总体为团状分布,且除紫椴外,其余树种不均匀分布的比例都要大于均匀分布的比例。由全林分整体来看,仍以随机分布所占比例最大,为33%,而绝对均匀和绝对不均匀分布所占比例分别只有12%和9%,整个林分的平均角尺度为0.48,整体呈随机分布。
4 结论与讨论
本研究未采用传统的大小比数来分析树种的优劣程度,而采用直观的胸径结构、树高结构来反映其林分水平的竞争状态,结果表明,作为优势树种的山杨在胸径和树高上都集中在较大径阶,表明无论在水平方向还是垂直方向上,其都具有显著竞争优势;与之相反,刺楸主要集中在较小径阶,因此在竞争中不占优势。
本研究采用树种混交度指标,从单木的角度描述林分树种的空间搭配及排列形式,天然山杨林在空间结构单元中主要以强度混交为主,处于中度和极强度混交的林木比例也比较大,说明天然山杨林整体上的混交状况良好,同种隔离程度较高。研究表明,各树种平均混交度依次是:紫椴(0.90)>黄桦(0.89)>大叶杨(0.86)>色木槭(0.82)>蒙古栎(0.79)>花曲柳(0.75)>白桦(0.71)>花楷槭(0.69)>刺楸(0.67)>千金榆(0.55)>山杨(0.45),不难看出,作为优势树种的山杨混交程度是最低的。全林分平均混交度达到0.65,说明天然山杨林混交程度较高,林分比较稳定。
天然山杨林内林木有的呈丛状聚集分布,可能经过自疏或人为干扰,成林后林木聚集程度有所下降,研究全林分平均角尺度发现,林分整体呈随机分布,且绝对均匀和绝对不均匀分布这2种极端情况所占比例非常小。就各树种而言,蒙古栎、黄桦、刺楸、花楷槭和千金榆呈均匀分布;山杨呈随机分布;大叶杨、色木槭、紫椴、花曲柳和白桦的平均角尺度均比0.517要稍大,说明它们总体为团状分布。
利用以空间结构单元为基础的树种混交度和角尺度等参数分析林分的空间结构,便于人们对森林结构有一个比较清晰的认识[16]。在调整森林结构时,应针对顶极或主要伴生树种的胸径和树高结构特征、角尺度、混交度来进行空间结构调整,使经营对象处于竞争优势或不受到挤压的威胁,使整个林分的格局趋于随机分布,群落生物多样性得到提高,林分群体均获得健康。
[参考文献]
[1] 闫东锋,何 静,郭 芳,等.宝天曼栎类天然次生林乔木层种间联结性研究 [J].西北林学院学报,2009,24(1):26-30.
Yan D F,He J,Guo F,et al.Interspecific association of main tree populations of oak natural secondary forest in Baotianman [J].Journal of Northwest Forestry University,2009,24(1):26-30.(in Chinese)
[2] 柴 勇,孟广涛,武 力,等.馨香玉兰所在群落主要树种的种间联结性 [J].西北林学院学报,2009,24(5):31-35.
Chai Y,Meng G T,Wu L,et al.Interspecific associations of main tree species in the communities with magnoliaod or atissima [J].Journal of Northwest Forestry University,2009,24(5):31-35.(in Chinese)
[3] 李明辉,何风华,刘 云,等.林分空间格局的研究方法 [J].生态科学,2003,22(1):43-50.
Li M H,He F H, Liu Y,et al.Analysis methods of stand spatial distribution pattern [J].Ecologic Science,2003,22 (1):43-50.( in Chinese)
[4] 胡艳波,惠刚盈.优化林分空间结构的森林经营方法探讨 [J].林业科学研究,2006,19(1):1-8.
Hu Y B,Hui G Y.A discussion on forest management method optimizing forest spatial structure [J].Forest Research,2006,19(1):1-8.(in Chinese)
[5] 惠刚盈,Von Gadow K,Albert M.角尺度:一个描述林木个体分布格局的结构参数 [J].林业科学,1999,35(1):37-42.
Hui G Y,Von Gadow K,Albert M.The neighbourhood pattern:A new structure parameter for describing distribution of forest tree position [J].Scientia Silvae Sinicae,1999,35(1):37-42.(in Chinese)
[6] 惠刚盈,Von Gadow K,Albert M.一个新的林分空间结构参数:大小比数 [J].林业科学研究,1999,35(1):1-6.
Hui G Y,Von Gadow K,Albert M.A new parameter for stand spatial structure:Neighbourhood comparison [J].Forest Research,1999,35(1):1-6.(in Chinese)
[7] 惠刚盈,胡艳波.混交林树种空间隔离程度表达方式的研究 [J].林业科学研究,2001,14(1):177-181.
Hui G Y,Hu Y B.Measuring species spatial isolation in mixed forests [J].Forest Research,2001,14(1):177-181.(in Chinese)
[8] 覃 林,谭 玲,罗应华,等.广西大明山云贵山茉莉林的空间结构 [J].中南林业调查规划,2008,27(2):66-69.
Qin L,Tan L,Luo Y H,et al.The spatial structure ofHoudendronbiaristatumforest in Daming Mountain Nature Reserve,Guangxi [J].Central South Forest Inventory and Planning,2008,27(2):66-69.(in Chinese)
[9] 李明辉,何风华,刘 云,等.林分空间格局的研究方法 [J].生态科学,2003,22(1):43-50.
Li M H,He F H,Liu Y,et al.Analysis methods of stand spatial distribution pattern [J].Ecologic Science,2003,22(1):43-50.(in Chinese)
[10] Gadow K V.Continuous cover forest assessment,analysis scenarios [C]//International IUFRO Conference.Goettingen,Germany:[s.n.],2001:19-21.
[11] 吴富桢.测树学 [M].北京:中国林业出版社,1992.
Wu F Z.Measurement of trees [M].Beijing:China Forestry Publishing House,1992.(in Chinese)
[12] 惠刚盈,Gasow K V,胡艳波.林分空间结构参数角尺度的标准角选择 [J].林业科学研究,2004,17(6):687-692.
Hui G Y,Gasow K V,Hu Y B.The selection of standard angle of the neighbourhood pattern,a stand spatial structure parameter [J].Forest Research,2004,17(6):687-692.(in Chinese)
[13] Gadow K V,Hui G Y.Characterizing forest spatial structure and diversity [C]//Sustainable Forestry in Temperate Regions.Sweden:University of Lund,2002.
[14] 胡艳波,惠刚盈,戚继忠,等.天然红松阔叶林的空间结构分析 [J].林业科学研究,2003,16(5):523-530.
Hu Y B,Hui G Y,Qi J Z,et al.Analysis of the spatial structure of natural Korean pine broad leaved forest [J].Forest Research,2003,16(5):523-530.(in Chinese)
[15] 邓送求,闫家锋.南京紫金山枫香风景林空间结构分析 [J].南京林业大学学报:自然科学版,2010,34(4):117-122.
Deng S Q,Yan J F.Spatial structure of scenic forest ofLiquidamabarformosanain Nanjing Zijin Mountain [J].Journal of Nanjing Forestry University:Natural Sciences,2010,34(4):117-122.(in Chinese)
[16] Dungan J L,Perry J N,Dale R T,et al.A balanced view of scale in spatial statistical analysis [J].Ecography,2002,25:626-640.
猜你喜欢
现代农村科技(2022年5期)2022-11-18
阿来研究(2020年1期)2020-10-28
防护林科技(2020年6期)2020-08-12
绿色科技(2019年6期)2019-04-12
绿色科技(2019年6期)2019-04-12
现代农业研究(2017年11期)2018-01-12
现代园艺(2017年20期)2017-02-03
新世纪水泥导报(2016年1期)2016-07-01
农业与技术(2016年6期)2016-05-14
中央社会主义学院学报(2016年2期)2016-05-04
