
三疣梭子蟹两种养殖模式浮游动物调查
2014-03-23 02:24毛欣欣蒋霞敏王春琳伊祥华
生物学杂志 2014年4期
毛欣欣, 蒋霞敏, 王春琳, 伊祥华
(1. 宁波大学 海洋学院, 浙江 宁波 315211; 2. 象山双华生态养殖有限公司,浙江 宁波 315211)
三疣梭子蟹(Portunustrituberculatus)是中国池塘养殖的重要海产经济蟹类。近年来,梭子蟹的养殖模式呈现多样化的态势[1],主要有单养、虾贝混养等[2]。单养不但易发病,而且池塘利用率低,不利于三疣梭子蟹养殖业的可持续发展;多品种立体化混养可以提高产量,防止水污染[3-6]。目前由于养殖模式简单、落后,养殖产量始终徘徊在450~600 kg/hm2,养殖效益提高不显著[7]。浮游动物是水生生态系统的初级消费者,是水体生态系统食物链中一个重要的环节[8],养殖水体的浮游动物不仅可提供基础饵料,也可对水体生态系统的结构和功能起到重要的调节作用。但目前有关浮游动物的种类、数量与三疣梭子蟹的生长和产量的关系未见报道。
本文通过对两种养殖模式三疣梭子蟹池塘浮游动物种类组成及数量变动的调查,探讨其与环境因子的关系,旨在深入认识三疣梭子蟹养殖池塘浮游动物的生态特征,为健康养殖提供基础资料,为合理制定高产的养殖模式提供参考依据。
1 材料和方法
1.1 两种养殖模式
试验于2012年6月—12月在宁波市象山县东盛水产养殖基地池塘进行,选择两种养殖模式,每种养殖模式设3个池塘。模式Ⅰ面积2亩;土塘、四周水泥、底铺20目网、周围铺2 m砂;三疣梭子蟹与日本对虾、黑鲷、菲律宾蛤仔混养;底充氧增氧。模式Ⅱ面积5亩;土塘、四周铺黑膜;三疣梭子蟹与日本对虾、黑鲷混养;增氧机增氧。两种模式都放养三疣梭子蟹35尾/m2,2期苗。日本对虾12尾/m2,黑鲷60尾/m2,规格一致。模式Ⅰ放养菲律宾蛤仔88个/m2。每天定时投喂饵料小杂鱼,每个池塘的量相当。
1.2 浮游动物样品的采集及处理
实验期间每月采集浮游动物1次,采样时间一般为上午10:00—12:00,采样点位于池塘4个角落离开1 m处。调查研究方法按照《海洋生物调查规范》进行。大型浮游动物定性水样用13#浮游生物网在采水点“∞”字型拖10 min,定量水样用有机玻璃采水器采集混合样10 L。所采浮游动物用甲醛溶液固定。定量水样带回实验室沉淀48 h,浓缩至20 mL,吸取1.0 mL置于浮游动物计数框中在10×10倍显微镜下全片计数,原生动物和小型轮虫及无节幼体的定性采用浮游植物定性样品,定量采用浮游植物定量的浓缩水样进一步浓缩至20 mL,再进行计数。每瓶标本计数3次,取其平均值。
1.3 多样性数据分析
优势度Y按照徐兆礼[9]采用如下公式计算:
Y=Pi×fi(Pi=ni/N)
式中,ni为丰度,fi是出现率,N为总丰度。取优势度Y≥0.02的种类为优势种。
多样性指数主要参照文献[10],计算公式如下:
马格利夫(Margalef)指数:d=(S-1)/lnN
香农(Shannon-Wiener)指数:H′ =-∑Pi×log2Pi(Pi=ni/N)
均匀度(Pielou)指数:J=H′/Hmax,Hmax=log2S
式中:S种类数;N总密度;ni第i种的密度。
1.4 数据处理
采用SPSS 17.0软件对试验数据进行相关分析。
2 结果
2.1 水质指标
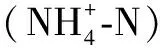
2.2 种类组成
两个模式中鉴定出的浮游动物共13属19种。其中,原生动物3种,占总数的15.8%,隶属于1纲3属;轮虫5种,占总数的26.3%,隶属于2科2属;枝角类3种,占总数的15.8%,隶属于2科2属;桡足类8种,占总数的42.1%,隶属于4科6属。每月浮游动物的种类组成见图1。由图1可以看出,6、7、8和12月4个月份浮游动物种数均为模式Ⅰ>模式Ⅱ,9月模式Ⅰ<模式Ⅱ,10和11月模式Ⅰ=模式Ⅱ。每个月份2种模式桡足类都占绝对优势。

图1 两种混养模式不同月份浮游动物种类组成
2.3 优势种群
不同月份两种养殖模式的养殖水体中出现的优势种差异显著(表1)。由表1可以看出,养殖期内桡足类无节幼体在6—11月一直是模式Ⅰ和模式Ⅱ优势种群,在12月模式Ⅰ中仍为优势种,模式Ⅱ中被桡足幼体所替代。而模式Ⅰ6月和10月出现了轮虫优势种壶状臂尾轮虫Brachionusurceus、褶皱臂尾轮虫Brachionusplicatilis,11月出现了枝角类优势种蚤状蚤Daphniapulex。7—12月中华拟铃虫Leprotintinnus都是模式Ⅰ优势种。大同长腹剑水蚤Oithonidaesimilis是9月、12月模式Ⅱ优势种,湖泊美丽猛水蚤Nitocralacustris11月、12月模式Ⅰ优势种。模式Ⅱ优势种相对较少。
2.4 密度和生物量
从调查结果可见,两种养殖模式池塘浮游动物密度较高,生物量较大。由于浮游动物个体差别较大,计数方法不同,在计算数量时,将其分为大型浮游动物(包括轮虫、枝角类、桡足类)和原生动物两部分。养殖周期中,两种模式浮游动物的密度变化范围为47~958 ind./L,平均密度模式Ⅰ(370 ind./L)>模式Ⅱ(209 ind./L)。模式Ⅰ6—8月先降后升,8月份达到最大值(958 ind./L),以后又逐渐下降。模式Ⅱ浮游动物密度6月最大(543 ind/L),以后逐月呈波浪状高低起伏变化。

表1 两种混养模式不同月份的优势种
“+”为优势种;“-”为非优势种。

表2 两种模式不同月份浮游动物密度(ind./L)
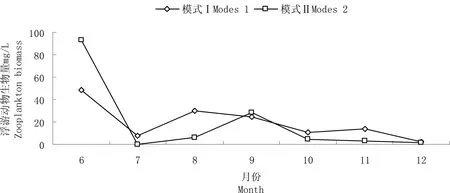
图2不同混养模式浮游动物生物量随月份的变化
Fig 2 Zooplankton biomass of different months in two polyculture modes
两种养殖模式的浮游动物生物量变化范围为0.182~93.076 mg/L,平均生物量模式Ⅱ(18.102 mg/L)>模式Ⅰ(17.974 mg/L)。由图2可以看出,两种模式在6月份生物量最大。6—8月,两种模式都呈现先降后升的趋势,模式Ⅱ生物量差异显著。9—12月,模式Ⅰ先降后升再降,模式Ⅱ一直在下降。
2.5 多样性指数
养殖期间中,两种模式的浮游动物多样性指数见表3。由表3可知,两种模式的Margalef指数、Shannon-Wiener指数、Pielou均匀度指数均呈现波动变化,变化趋势基本一致,且均值都是模式Ⅰ>模式Ⅱ。Margalef指数变化范围为0.22~0.81;Shannon-Wiener指数变化范围为1.28~3.03;Pielou均匀度指数变化范围为0.54~0.96。

表3 两种混养模式浮游动物的多样性指数
3 讨论
3.1 两种模式浮游动物群落结构特征
本次调查两种模式共鉴定出浮游动物19种,其中,原生动物3种,占总数的15.8%;轮虫5种,占总数的26.3%;枝角类3种,占总数的15.8%;桡足类8种,占总数的42.1%。模式Ⅰ种类数6、8、11月最多(9种),9月最少(5种);模式Ⅱ种类数11月最多(9种),7月最少(3种)。平均种类数模式Ⅰ>模式Ⅱ。桡足类在每个月份都占据了绝对优势形成优势种群。两种模式优势种有所区别,壶状臂尾轮虫、褶皱臂尾轮虫、蚤状蚤、湖泊美丽猛水蚤都仅在模式Ⅰ成为优势种。密度和生物量均为模式Ⅰ>模式Ⅱ,且随月份变化显著。多样性指数均值也是模式Ⅰ>模式Ⅱ。
比较之下,模式Ⅰ浮游动物群落结构较好。究其原因,可能是混养种类与增氧方式的不同引起的。蟹虾鱼贝混养,蟹、虾食性相近,但蟹类能捕食一些虾类不能利用的大型饵料,而虾类又能吃一些蟹类不能摄取的细小生物和饵料硝屑,这样就减少了残饵污染水质[11];鱼能将活动迟缓的病虾、病蟹捕食,杜绝了病害的蔓延;同时利用贝类的滤食特点,充分利用蟹、虾残饵、浮游藻类、底栖生物,净化了水质[12-14]。底充氧利用管道对水体充氧,不会把水体搅浑,使水体成富氧状态,能迅速有效地分解水中的有害物,同时促使池塘水体上下翻腾避免了盐跃层和氧跃层,维持良好的水环境[15, 16]。模式Ⅰ水质好,浮游动物群落结构丰富。
3.2 影响浮游动物群落结构的环境因子分析
浮游动物的生命周期较长,群落相对稳定,对环境的变化敏感,是反映海洋环境变化的理想研究对象[17]。由表4可以看出,浮游动物密度、生物量与温度、溶氧呈现正相关,与盐度、pH值、透明度呈现负相关,说明随着水温的升高和溶氧的增加,浮游动物群落多样性有增大的趋势,而盐度、pH值、透明度的增加,群落多样性有减小的趋势。多样性指数与溶氧、温度呈现正相关,与透明度、盐度、pH值呈现负相关。模式Ⅰ中浮游动物密度与温度呈显著正相关,与pH值呈显著负相关,生物量与盐度呈显著负相关。模式Ⅱ中浮游动物密度与溶氧呈显著正相关,与盐度、pH值呈显著负相关,其中生物量与盐度呈高度显著负相关。多样性指数Shannon-Wiener指数和Pielou均匀度指数仅在模式Ⅰ出现与温度呈显著正相关,与透明度呈显著负相关,而Margalef指数仅在模式Ⅱ中出现与总磷呈显著正相关,其中Pielou均匀度指数与温度呈现高度显著正相关。
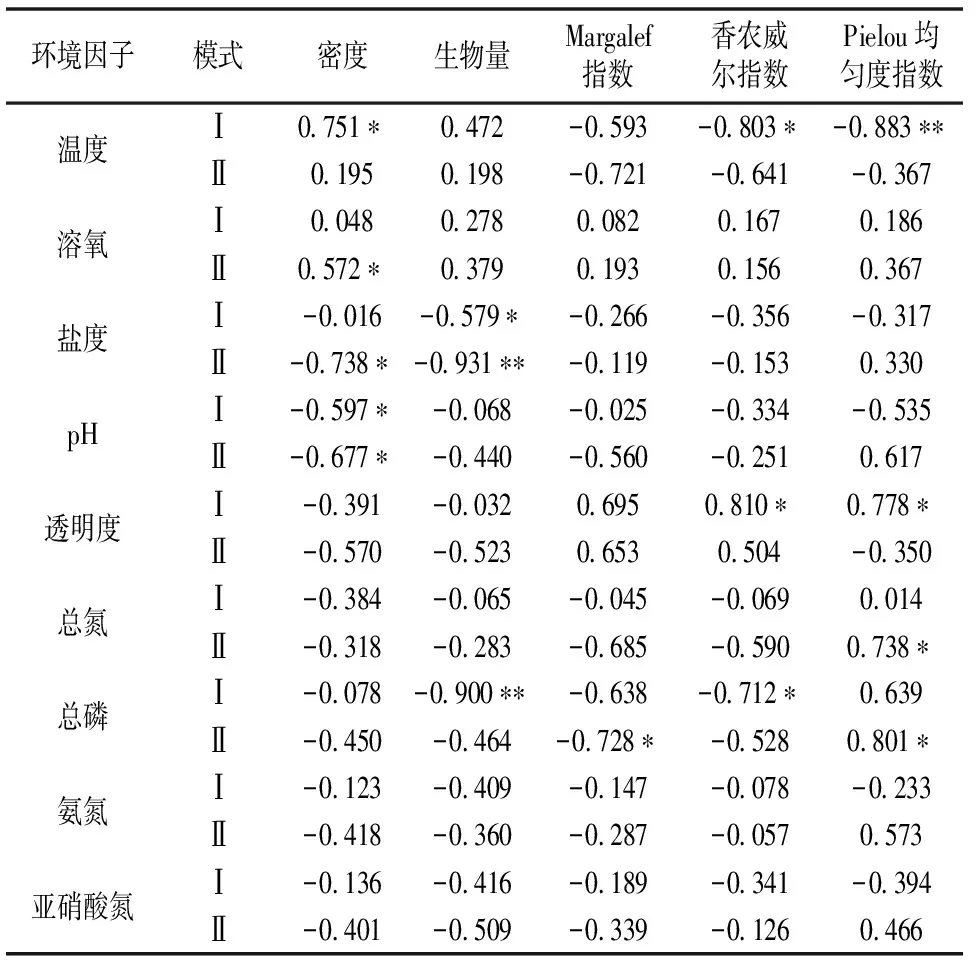
表4 浮游动物密度、生物量及多样性指数与环境因子的相关系数
*—P<0.05; **—P<0.01。
营养盐是浮游动物生存的基础条件,营养盐的变化也会影响浮游动物的数量与分布。在某种意义上说,正常的营养盐浓度是维持浮游生物一定结构和丰度的物质基础和能量来源[18]。表4中,浮游动物密度、生物量、多样性指数基本都与营养盐含量呈现负相关,说明随着营养盐溶度升高,浮游动物密度、生物量、多样性指数下降,这可能是浮游动物生态群落发生了变化,有生态异常和不稳定趋势,这也是对环境信息的一种反馈[19]。
参考文献:
[1]陈 芳, 孙 忠, 王跃斌. 三疣梭子蟹不同养殖模式池塘夏季溶解氧变化特征的研究[J]. 浙江海洋学院学报:自然科学版, 2011, 30(3): 221-225.
[2]张达云. 三疣梭子蟹池塘生态养殖技术[J]. 现代农业科技, 2010(10): 317-318.
[3]原永党, 孙本腾, 毕复永, 等. 梭子蟹池塘沉箱式单养试验[J]. 海洋湖沼通报, 2002(3): 80-84.
[4]张兴国. 脊尾白虾与梭子蟹、贝类混养技术[J]. 齐鲁渔业, 2008, 25(4): 35-36.
[5]王兴强, 曹 梅, 阎斌伦, 等. 三疣梭子蟹综合养殖技术[J]. 水产科学, 2009, 28(2): 105-108.
[6]石志洲. 三疣梭子蟹池塘养殖技术[J]. 海洋渔业,1999(3): 129-130.
[7]施祥元, 伊祥华, 郑凯宏, 等. 三疣梭子蟹底充氧养殖模式的研究[J]. 河北渔业, 2008(3): 13-15.
[8]陈 亮, 刘 一, 禹 娜, 等. 分水江水库浮游动物群落结构的初步研究及水质评价[J]. 华东师范大学学报:自然科学版, 2010(6): 72-82.
[9]徐兆礼. 东海亚强真哲水蚤种群生态特征[J]. 生态学报, 2006, 26(4): 1151-1158.
[10]孙儒泳, 李 博, 诸葛阳, 等. 普通生态学[M]. 北京: 高等教育出版社, 1993: 136-137.
[11]胡则辉, 徐君卓, 石建高. 浙江沿海三疣梭子蟹的养殖模式[J]. 现代渔业信息, 2011, 26(3): 3-5.
[12]林庆贵. 三疣梭子蟹、日本对虾、花蛤混养技术[J]. 齐鲁渔业, 2010, 7(4): 35-36.
[13]陈金仙. 三疣梭子蟹、日本对虾、花蛤混养技术[J]. 中国水产, 2008(12): 52-53.
[14]欧海金. 梭子蟹、脊尾白虾、贝类的立体生态混养技术[J]. 新农村, 2010: 34-35.
[15]马亚梅. 虾蟹养殖中池塘水质与底质的调控[J]. 渔业致富指南, 2011(20): 25-27.
[16]蔡建中, 李洪进, 唐玉华. 鱼虾蟹池塘微孔增氧高产高效养殖技术初探[J]. 渔业致富指南, 2011(1): 55-57.
[17]张海生, 宁修仁, 乐凤凤, 等. 三门湾沉积记录中长链烯酮不饱和指数-海表温度和浮游动物群落对厄尔尼诺/拉尼娜的响应[J]. 海洋学报, 2010, 32(3): 40-50.
[18]傅彩萍. “底充氧”对养殖池塘水质和浮游生物群落结构的影响[D]. 宁波:宁波大学, 2010.
[19]孙晓庆, 董树刚, 汤志宏. 营养盐和光照对浮游植物群落结构的影响[J]. 南方水产, 2008, 4(1): 1-9.
猜你喜欢
今日农业(2020年19期)2020-12-14
科学养鱼(2020年10期)2020-11-23
课外语文·中(2020年2期)2020-03-24
课外语文(2020年5期)2020-03-14
知识窗(2019年4期)2019-04-26
海峡姐妹(2018年11期)2018-12-19
北京广播电视报(2018年39期)2018-11-23
现代营销·经营版(2017年3期)2017-05-17
作文周刊·小学五年级版(2016年1期)2016-06-29
中国科技信息(2015年2期)2015-11-16
