
杭州湾滩涂湿地植被主要植物的光能利用率和光化学指数的相关性研究
2014-03-20 01:13:34王景旭丁丽霞
杭州师范大学学报(自然科学版) 2014年6期
王景旭,丁丽霞,程 乾
(1.浙江农林大学浙江省森林生态系统碳循环与固碳减排重点实验室,浙江 临安311300;2.浙江工商大学旅游与城市管理学院,浙江 杭州310018)
0 引 言
光能利用率LUE(Light Use Efficiency)是估算净初级生产力NPP(Net Primary Productivity)的一个关键变量,用来表征植物截获/吸收太阳能转换成有机干物质的效率[1-2].在众多估算NPP和GPP(总初级生产力,Gross Primary Productivity)的模型中,光能利用率LUE模型应用最广.LUE模型具有机理明确、计算简单、参数较少和易与遥感数据结合等优点,已成为估算GPP和NPP的主要方法[3-4].与其他模型对比研究表明,基于LUE的遥感模型模拟结果偏离较大,而且LUE是主要误差[4],因此获取可靠精度的LUE对研究能量的分布以及全球环境的变化有着重要的意义[5].近年来湿地所具有的多种功能和价值逐渐为人们所认知.湿地生态系统中的能量贮存和流动是其维持和发展的基础.杭州湾滨海滩涂湿地是中国滨海湿地的南北过渡带,在维持区域生态平衡、提供珍稀动植物栖息地和保护生物多样性等方面具有非常重要的作用[6].
近些年来的研究表明,在叶片[7]、冠层[8]和景观尺度[9]上,基于遥感的光化学指数PRI(Photochemical Reflectance Index)来无损估计实际LUE是完全可行的.Gamon把PRI定义为531和570 nm 处反射率的归一化植被指数[10-12].这两个波段位置的反射率受到叶黄质循环的影响并和叶片的光能利用率有密切相关.因此PRI为估算叶片的光能利用率提供了一种新的方法.植物在长期进化中产生了一套自我保护机制,来避免过剩光能对光合机构造成破坏,其中叶黄质循环被认为是热耗散的重要保护机制之一[13-15]:当入射光强过多,超过植物光合作用所需时,就会引起叶黄质从环氧化状态变为脱环氧化状态,来散失热量以保护光合机构;反之,叶黄质则朝相反的方向进行,形成循环来提高光合作用效率.所以叶黄质循环能很好地表现LUE的变化.而在很多研究中,叶黄质循环和531 nm 的反射率有很强的相关性[16],这就使得PRI能在自然条件下很好地来估算光合利用率.
吴统贵等[17]以杭州湾不同年份形成的滩涂湿地植被群落为研究对象,指出该区域植被群落演替序列为海三棱藨草(Scirpusmariqueter)群落→芦苇(Phragmitesaustralis)群落→柽柳(Tamarixchinensis)+芦苇群落→旱柳(Salixmatsudana)+白茅(Imperatacylindrica)群落.对芦苇、柽柳和旱柳这3种植物的研究能很好地表现出杭州湾湿地的状态.因此建立柽柳、芦苇和旱柳LUE和PRI的相关关系,可以结合遥感图像,开展大面积的杭州湾湿地生产力的估算,为人们提高湿地生产力、保护生态环境提供科学对策.
1 研究区和数据获取
1.1 研究区
研究区(121°08′42″E,30°18′40″N)位于浙江省慈溪市杭州湾湿地,多年平均气温为16.0 ℃,全年1月份最冷,平均气温为3.8℃,7月份最热,平均气温28.2℃,全年平均日照时数约为2 038.4 h,无霜期约为244 d.多年平均降水量为1 344.7 mm.同时,滩涂湿地受潮水影响较大,该区域潮流属不正归日潮流,为往复流性质,月平均潮位随季节变化,9月份最高,1月份最低,年变幅0.5 m.土壤为长江水和浙江入海河流输沙和海底掀沙淤积而成,属于滨海盐土带.在植被区划上该区属北亚热带常绿阔叶林北部亚地带[17].
本文选取植物叶片处于成熟期的10月份,选择晴朗少云的天气进行数据测定.叶面积指数LAI(Leaf Area Index)作为影响PRI的一个重要因子,选择低LAI植物柽柳、中LAI植物芦苇以及高LAI植物旱柳作为观测研究对象,可以说明不同LAI对PRI和LUE相关性的影响.
1.2 观测方法
1.2.1 冠层光谱的测定
3种植被的冠层光谱采用美国Analytical Spectral Device(ASD)公司生产的Field-Spec Pro FR2500型背挂式野外高光谱辐射仪测量.测量波段为350~2 500 nm,其中350~1 000 nm 间的光谱采样间隔为1.4 nm.冠层光谱的测定选在晴朗、少云、风速较小时进行.测定时间为2013年10月11日8:30-16:00,测量时传感器探头垂直向下,光谱仪视场角为25°,距冠层垂直高度为0.5 m.每个观测点记录10个光谱数据,以其平均值作为该观测点的光谱反射值.测量过程中,及时地对每次观测进行标准白板校正(白板反射率为0.99),因此所得目标物光谱为相对反射率.
PRI的计算依据Gamon等据向日葵生化特性短期变化提出的PRI计算公式[9],本文为计算方便,把公式稍作修改如下:
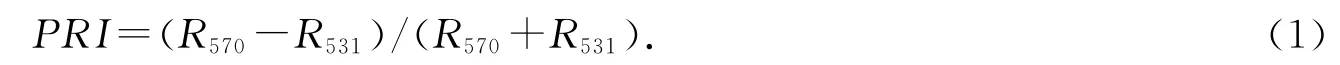
其中,R531和R570分别表示531和570 nm 处的冠层反射率.R531能敏感地反映出叶黄素循环组分的变化.选取R570作为参考是为了消除其他色素的影响.
试验分别测得每种植被6组对应的PRI和LUE数据,并进行PRI和LUE的相关分析.
1.2.2 冠层光合作用数据的获取
与光谱测量同步,植物光合作用采用英国ADC 公司的LCPro-SD 便携式光合仪测定.取光谱测量的10株植物作为实验样本,测量植株的上、中、下3层具有代表性的叶各1片,每片叶取5个重复值,以3片叶的平均值来代表整个植被冠层光合作用[18].LCPro-SD光合作用仪可以测得植物叶片的光合有效辐射、光合速率、蒸腾速率、气孔导度等光合作用参数.
LUE定义为冠层截取的每摩尔光合有效辐射中碳的固定量,即在一定时间内,植物吸收光能的效率.LUE的计算公式如下[19]:

其中Pn表示CO2的固定速率,即测定的叶片净光合速率;PPFD(Photosynthetic Photon Flux Density)代表冠层截获的光合有效辐射.
试验测定3个植被各7组主要生理指标和环境因子数据,并用SPSS进行相关分析.
1.2.3LAI测定
叶面积指数LAI可以通过WinSCANOPY 冠层分析仪(加拿大Regent Instruments公司产品,包括Minolta Dimage Xt数码相机和外接Nikon FC2E8鱼眼镜头)获取,对每株植物进行半球图像采集,所得半球影响的数码照片通过WinSCANOPY For Hemispherical Image Analysis分析软件进行分析,获得每株植被的LAI.
2 结果分析
2.1 LUE 对光照的日变化响应
光照强度是影响植物光合作用最重要的因素.由图1可知,光合有效辐射PAR(Photosynthetically Active Radiation)在8:30时已经达到了864.37μmol·m-2·s-1,在11:30左右达到最大值,然后随着时间推移逐渐降低.其他的环境因子如温度(T)也随之变化.温度在8:30的时候达到了33.36 ℃,之后随着光强的增大而增大.但是到达顶峰的时间迟于光照,在12:30到达39℃后降低.由于植物夜间进行呼吸作用,释放CO2,所以早晨大气CO2浓度较高.随着光照的不断增加,植物光合作用的不断增强,CO2浓度也随之不断降低.
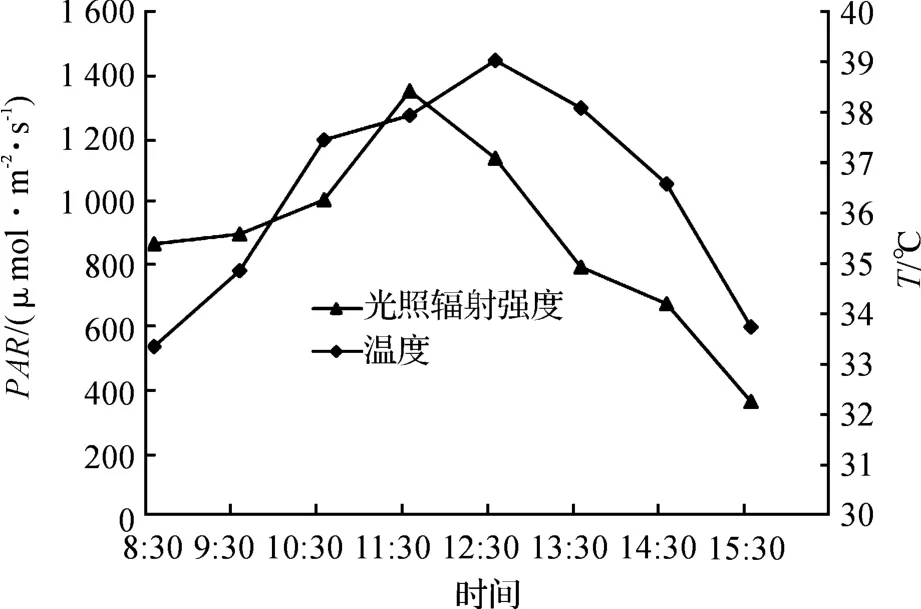
图1 环境因子的日变化(2013-10-11)Fig.1 The diurnal variation of the environmental factors
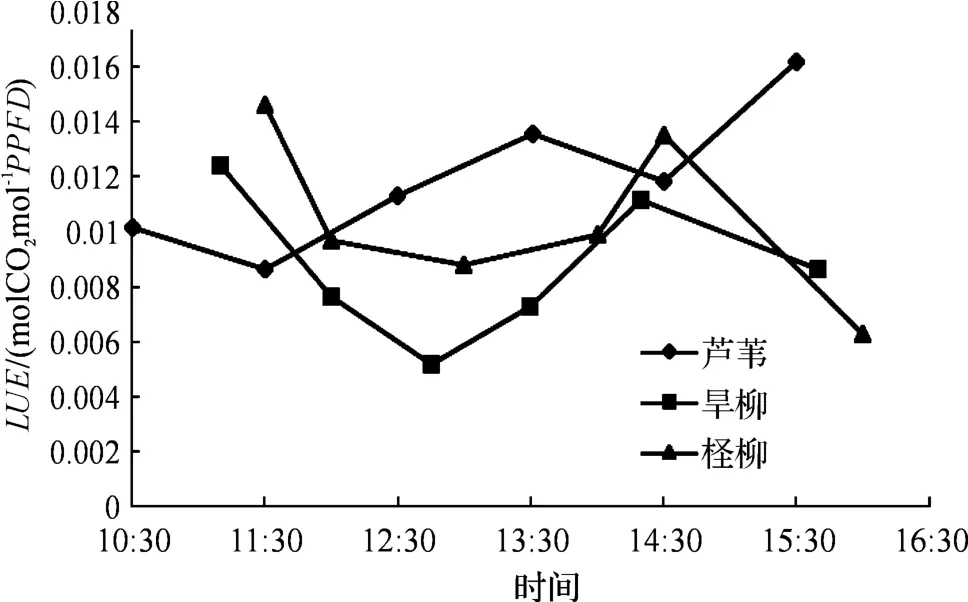
图2 3种植物的LUE 日变化曲线(2013-10-11)Fig.2 The daily change curve of LUE of three kinds of plants
由图2中可知:柽柳和旱柳的LUE曲线都有一个低谷,二者的LUE变化趋势相同,而芦苇跟二者的变化差别很大.柽柳和旱柳属于双子叶乔木,芦苇是单子叶草本植物,三者的叶片结构不同.在相同环境下,三者的光能利用率对随时间变化的光强响应是不同的.旱柳最早达到LUE的最大值,在10:30的时候达到0.012 44;其次是柽柳,在11:30达到0.014 62的最大值;芦苇达到最大值的时间最晚,LUE一直呈阶梯状上升趋势,在15:30达到0.016 19.光照强度达到最大之前,柽柳LUE>芦苇LUE>旱柳LUE,当光照逐渐减小,芦苇LUE>柽柳LUE>旱柳LUE.
2.2 净光合速率和相关生理生态因子的日变化
由图3中3种植物的生理指标可以看出,随着周围环境的变化,植物的净光合速率(Pn)、胞间CO2浓度(Ci)和气孔导度(Gs)也会发生相应的日变化.净光合速率是植物最重要的生理指标,反映了植物同化CO2的能力.由图3中可以看出,净光合速率因植被的不同而呈现不同的变化趋势.3种湿地植被的净光合速率在11:30后的大小顺序为:芦苇>柽柳>旱柳,而且芦苇比其它二者更晚达到最顶峰.三者受到的有效光合辐射随着时间的变化趋势一致,都在12:00左右达到最大.
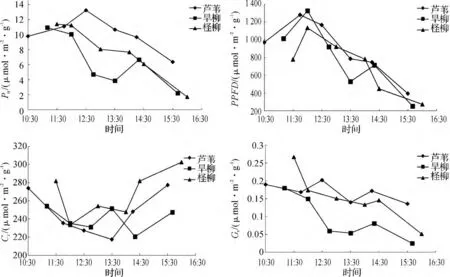
图3 芦苇、旱柳和柽柳叶片的生理指标日变化曲线(2013-10-11)Fig.3 The diurnal variation curve of blade physiological indexes of three kinds of plants
芦苇属于阔叶的草本植物,旱柳和柽柳属于双子叶乔木,芦苇和后两者的叶片结构和冠层完全不同,因此它们的生理指标变化趋势不同.由图3可以看出,芦苇和旱柳、柽柳的Gs和Gi变化曲线完全不同.
2.3 光合作用生理指标与环境因子的相关性分析
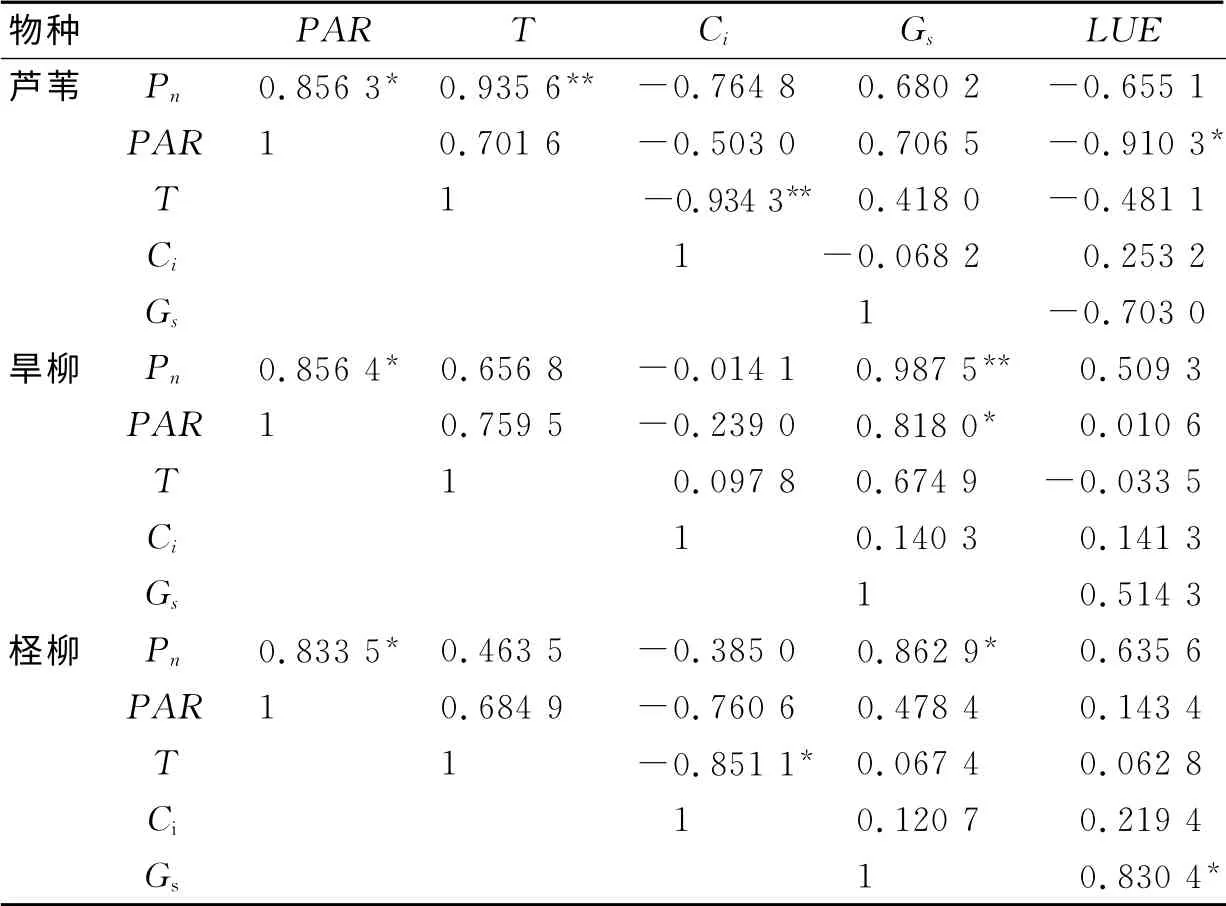
表1 净光合速率和主要生理生态因子间的相关关系Tab.1 Correlation between net photosynthetic rate and the main ecophysiological factors
植物的光合作用跟周围的生长环境有着复杂的关系,而且植物的各生理指标之间也存在着相互关系.对3种植被各7组主要生理指标和环境因子数据进行相关分析,可以发现不同植被的生理指标和生长环境之间存在的联系,结果见表1.
由表1 可以看出,3 种湿地植被的Pn与PAR、T、Gs都存在极显著的正相关;PAR与T、Gs呈正相关.植物自身的生理指标Gs和Ci不存在显著的相关关系;芦苇和旱柳的Gs随着T的增加而变大,柽柳的Gs则与T的变化无关;芦苇和柽柳的Ci与T、PAR和Pn存在极其显著的负相关,而旱柳的Ci与这3个因子无关;植被的光能利用率LUE与各因子有如下关系:芦苇的LUE与Pn、PAR和Gs呈负相关;旱柳和柽柳的LUE与Pn和Gs呈正相关;
2.4 PRI和LUE 的相关关系分析
芦苇、旱柳和柽柳的PRI指数随时间的变化如图4所示.对比图1、图2,在11:30之前,3种植被的PRI指数都随着光照增加而逐渐减小.在13:30-14:30,随着光照的降低,旱柳和柽柳的PRI指数反而升高,达到顶峰之后随光照降低而降低,而芦苇则升高.
图5反映的是3种湿地植物的PRI和LUE的相关关系.从图中可以看出,对于3种植物的冠层PRI对LUE的反应,判定PRI和LUE相关关系R2的大小顺序为:旱柳(0.956 5)>柽柳(0.951 5)>芦苇(0.642 5).这表明,在冠层尺度上,湿地植被的PRI对于植物的光能利用率LUE有很好的表达能力.图5可以明显地表现出LUE在整体水平上以芦苇最高,旱柳次之,柽柳最低.根据冠层分析仪得到的三者的冠层叶面积指数LAI分别为:旱柳(1.118)>柽柳(0.289)>芦苇(0.109 6).由此看出,植被在冠层尺度上的PRI与LUE关系受到植被叶面积指数LAI的直接影响.当LAI降低时,PRI指数和LUE都会受到外界因素的影响.因此,当LAI较低时,PRI和LUE的关系也会随之变小.冠层PRI和LUE的相关性在低LAI时较低,这是因为土壤PRI比冠层PRI低,在低LAI时,可见土壤较多,冠层PRI会被背景土壤的PRI拉低,所以在给定LUE下,高LAI的冠层PRI的相关性会比低LAI的相关性低[19-20].
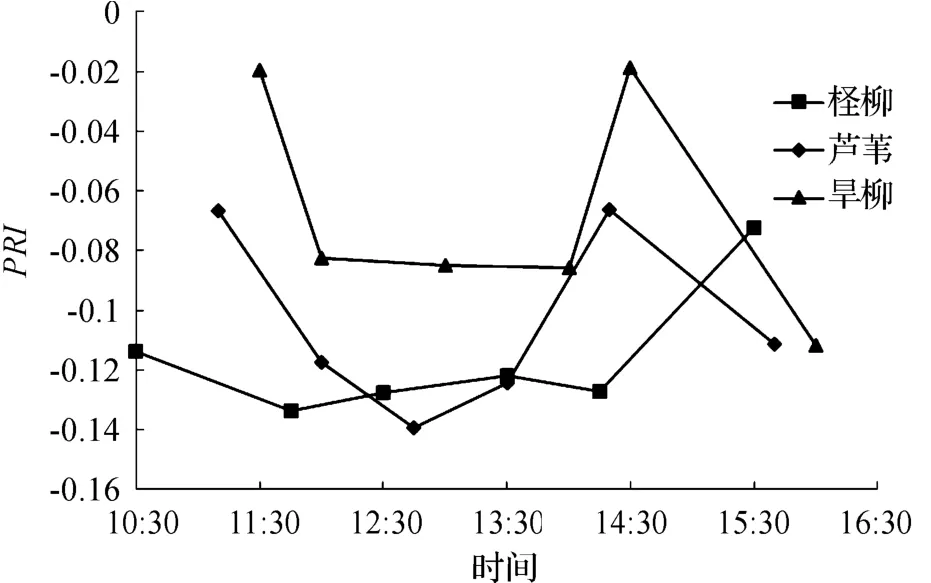
图4 3种植被PRI的日变化Fig.4 The diurnal variation of PRI of three kinds of plants
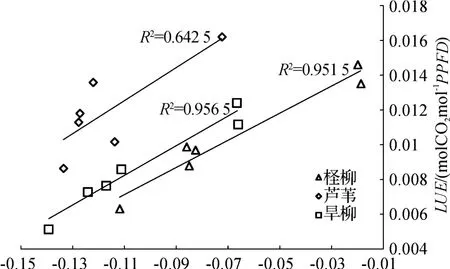
图5 3种湿地植被的PRI和LUE 的相关关系Fig.5 The correlation of PRI and LUE of three kinds of wetland vegetation respectively
3 结 论
本文研究了3种不同湿地植被的PRI和LUE的变化.结果表明,净光合速率因植被的不同而呈现不同的变化趋势,3种湿地植被的净光合速率在中午前后变化趋势为芦苇最大、柽柳次之、旱柳最小,而且芦苇比其它二者更晚达到最大,但三者的有效光合辐射随着时间的变化趋势一致,基本在中午12:00左右达到最大;3种湿地植被的Pn与PAR、T、Gs都存在极显著的正相关,PAR与T、Gs呈正相关,芦苇的LUE与Pn、PAR和Gs呈负相关,旱柳和柽柳的LUE与Pn和Gs呈正相关,与PAR无关;不同植被在冠层尺度上的PRI与LUE关系受到植被叶面积指数LAI的直接影响,3种湿地植被的PRI和LUE的相关性比较,芦苇最小,旱柳最大.
由于本文只是针对湿地3种植物开展了LUE和PRI的相关性及影响因素研究,因此所得结论还需要在更多不同类型植物中以及不同气候季节中开展研究加以验证.
[1]Monteith J L.Solar-radiation and productivity in tropical ecosystems[J].Journal of Applied Ecology,1972,9(3):747-766.
[2]Monteith J L,Moss C J.Climate and the efficiency of crop production in Britain[J].Philosophical Transactions of the Royal Society of London Series B:Biological Sciences,1977,281(980):277-294.
[3]Cramer W,Kicklighter D W,Bondeau A,etal.Comparing global models of terrestrial net primary productivity(NPP):overview and key results[J].Global Change Biology,1999,5(S1):1-15.
[4]Ruimy A,Kergoat L,Bondeau A,etal.Comparing global models of terrestrial net primary productivity(NPP):analysis of differences in light absorption and light-use efficiency[J].Glogal Change Biology,1999,5(1):56-64.
[5]Adams J M,Faure H,Faure-Denard L.Increases in terrestrial carbon storage from the last glacial maximum to the present[J].Nature,1990,348:711-714.
[6]王宪礼,李秀珍.湿地的国内外研究进展[J].生态学杂志,1997,16(1):58-63.
[7]Gamon J A,Serrano L,Surfus J S.The photochemical reflectance index:an optical indicator of photosynthetic radiation use efficiency across species,functional types,and nutrient levels[J].Oecologia,1997,112(4):492-501.
[8]Penuelas J,Llusia J,Pinol J,etal.Photochemical reflectance index and leaf photosynthetic radiation-use-efficiency assessment in mediterranean trees[J].International Journal of Remote Sensing,1997,18(13):2863-2868.
[9]Pentilde J,Inoue Y.Reflectance assessment of canopy CO2uptake[J].International Journal of Remote Sensing,2000,21(17):3353-3356.
[10]Gamon J A,Penuelas J,Field C B.A narrow-wave band spectral index that tracks diurnal changes in photosynthetic efficiency[J].Remote Sensing of Environment,1992,41(4):35-44.
[11]Justice C O,Vermote E,Townshend J R G,etal.The moderate resolution imaging spectroradiometer(MODIS):land remote sensing for global change research[J].IEEE Transactions on Geoscience and Remote Sensing,1998,36(4):1228-1249.
[12]Gamon J A,Field C B,Goulden M L,et.al.Relationships between NDVI,canopy structure,and photosynthesis in three Californian vegetation types[J].Ecological Applications,1995,5(1):28-41.
[13]Demmig-Adams B.Carotenoids and photoprotection in plants:a role for the xanthophyll zeaxanthin[J].Biochhim Biophys Acta,1990,1020(1):1-24.
[14]Gilmore A M.Mechanistic aspects of xanthophyll cycle-dependent photoprotection in higher plant chloroplasts and leaves[J].Physiol Plant,1997,99(1):197-209.
[15]Xu C G,Li L B,Kuang T Y.Photoprotection in chilling-sensitive and-resistant plants illuminated at a chilling temperature:role of the xanthophyll cycle in the protection against lumen acidification[J].Aust J Plant Physiol,2000,27(7):669-675.
[16]Gamon J A,Field C B,Bilger W,etal.Remote sensing of the xantophyll cycle and chlorophyll fluorescence in sunflower leaves and canopies[J].Oecologia,1990,85(1):1-7.
[17]吴统贵,吴明,萧江华.杭州湾滩涂湿地植被群落演替与物种多样性动态[J].生态学杂志,2008,27(8):1284-1289.
[18]Shrestha S,Brueck H,Asch F.Chlorophyll index,photochemical reflectance index and chlorophyll fluorescence measurements of rice leaves supplied with different N levels[J].Journal of Photochemistry and Photobiology B:Biology,2012,113:7-13.
[19]Long S P,Baker N R,Rains C A.Analyzing the responses of photosynthetic CO2assimilation to long-term elevation of atmospheric CO2concentration[J].Vegetatio,1993,14(104/105):33-45.
[20]Barton C V M,North P R J.Remote sensing of canopy light use efficiency using the photochemical reflectance index:model and sensitivity analysis[J].Remote Sensing of Environment,2001,78(3):264-273.
猜你喜欢
河北林业科技(2020年3期)2020-03-23 13:03:16
天津师范大学学报(自然科学版)(2019年4期)2019-09-17 00:59:10
山东林业科技(2018年4期)2018-09-01 08:15:20
作文周刊·小学四年级版(2018年12期)2018-04-10 12:07:56
花卉(2017年7期)2017-11-15 08:53:36
新课程·下旬(2017年7期)2017-08-14 11:12:50
花卉(2017年7期)2017-07-20 11:10:39
现代农业科技(2016年21期)2017-03-06 00:31:54
中国民族医药杂志(2016年8期)2016-05-09 07:51:05
西北林学院学报(2015年5期)2015-01-03 05:12:20
