
雷州马尾藻Sargassum leizhouense(Fucales,Phaeophyta)形态结构及分类研究*
2014-03-08 01:47:16赵素芬孙会强刘东超
海洋与湖沼 2014年2期
赵素芬 孙会强 刘东超①
(1.广东海洋大学水产学院 湛江 524025;2.广东海洋大学实验教学部 湛江 524088)
雷州马尾藻隶属于褐藻门(Phaeophyta),褐藻纲(Phaeophyceae),墨角藻目(Fucales),马尾藻科(Sargassaceae),马尾藻属(Sargassum)。马尾藻属最早由Agardh(1820)建立。目前,该属大约有 1000种(包括同物异名种),其中约 300多种被公认(Mattioet al,2011;赵素芬等,2012)。Agardh(1848,1889)根据藻体的叶片、气囊、生殖托等特征将该属分为5个亚属:叶枝亚属(Phyllotrichia)、裂叶亚属(Schizophycus)、反曲叶亚属(Bactrophycus)、节叶亚属(Arthrophycus)及真马尾藻亚属(Sargassum),其后该属的经典分类研究均是在此基础上修改并完善的。Womersley(1954)认为叶枝亚属(Phyllotrichia)和裂叶亚属(Schizophycus)的差异是分枝和气囊顶端小叶的发育程度,不足以作为亚属的划分标准,建议合并成叶枝亚属(Phyllotrichia)。曾呈奎等根据反曲叶亚属(Bactrophycus)种类生殖托既有单性、又有复合体的特点,建议将其与节叶亚属(Arthrophycus)合并(Tseng,1985;曾呈奎等,2000)。目前,马尾藻属的分类主要以叶枝亚属、反曲叶亚属、节叶亚属和真马尾藻亚属为参考依据。
由于富含膳食纤维、褐藻淀粉、矿物质和维生素等组分(王博等,2010),马尾藻一直是藻胶、食品、医药和饲料等领域的重要原料(李来好等,1997;刘东超等,2006a;Yeet al,2009)。近几年来,随着浅海马尾藻的过度采集,全球环境气候变化,以及潮间带海藻栖息环境的改变,导致马尾藻资源量急剧下降(Sunet al,2010),因此,大力开展马尾藻的人工繁育及增养殖尤为迫切(刘东超等,2006b;Xieet al,2013)。雷州马尾藻是雷州半岛潮间带常见的一种典型褐藻(曾呈奎等,1959,2000),由于该马尾藻藻体大,被用作鲍鱼饵料之一,并被大量采集作为饲料等工业原料,近五年其资源量明显减少(赵素芬等,2006),因此,迫切需要开展该马尾藻的形态学研究、人工繁育及增养殖工作,以改善浅海生态环境,构建藻场,为渔业资源的修复发挥重要作用。
本研究旨在通过海藻采集,结合藻体形态特征观察及记录,对我国雷州马尾藻进行形态特征、内部结构定量描述,重新对该马尾藻形态特征与分类地位进行研究,为其他马尾藻分类提供方法借鉴,也为开展雷州马尾藻的人工繁育、养殖及加工利用提供基础。
1 材料与方法
1.1 材料
雷州马尾藻取自湛江徐闻北港(20°13.855′N,110°6.725′E),在退潮时间隔 1—5m 随机采集 30—50棵,生长繁盛期与生殖期藻体分别采自 2010年 3月和2011年4月。采集后的藻体可直接用于数据测定和显微观测,或者暂养保存。实验室暂养条件为:海水温度20°C,海水盐度33,光照强度40—60 μmol/(m2•s),光照周期12L︰12D,充气培养。
1.2 方法
1.2.1 外部形态观测 将完整的藻体平均分为 3段,分别称基部、中部和顶端。肉眼观察并结合标尺测量,每组数据皆采用30—50个样本进行统计分析,主要记录马尾藻的藻体颜色,固着器、主干、初生分枝、次生分枝、叶片、生殖托和气囊等的形状与大小,分枝、生殖托和气囊的生长与分布特征,共计 20多个指标。
1.2.2 内部结构观测 从生长藻体中随机取10棵,从其中取30个次生分枝,切下顶端10cm 小段,进行徒手切片,用L1100A型生物电光显微镜观察藻体的内部结构;分别取10个发育早期(雌托长(2.71±0.70)mm,雄托长(3.68±1.21)mm)、中期(雌托长(4.57±0.80)mm,雄 托 长(6.64±1.85)mm)和 晚 期(雌 托 长(5.78± 1.27)mm,雄托长(15.71±4.62)mm)的雌、雄生殖托序,从中分别取30—50个生殖托,进行切片,然后在光学显微镜(OLYMPUS BH-2)下观察,记录藻体生殖托顶端1cm内的横切面结构特征。
1.2.3 数据分析 实验数据用SPSS 13.0数据处理系统进行统计分析,结果以平均值±标准误差表示。
1.2.4 形态与分类分析 根据雷州马尾藻的外部形态与内部结构特征进行统计分析,参照马尾藻属海藻的分类检索表(曾呈奎,2000),对雷州马尾藻的分类特征进行对比检验,对其分类地位进行修订。
2 结果
2.1 外部形态特征
2.1.1 生长期 藻体黄褐色至褐色,多生长在低潮带及潮下带1—2m深处(图1),藻体长达3m以上,由固着器、主干、初生分枝、球芽、叶片、次生分枝和气囊组成(图2)。固着器盘状。主干圆柱形,表面瘤状。初生分枝数条,由主干顶部长出,呈辐射状排列,下部(约占藻体长度的 2/3)扁压或扁平,上部(约占藻体长度的1/3)圆柱形或亚圆柱形,表面光滑。初生分枝基部分布有 2—5个椭圆形球芽,表面光滑。叶片长披针形,有一个短柄,基部略楔形,顶端尖细。叶上中肋明显,并延伸至叶尖,中肋两侧多毛窝,叶片边缘全缘或有锯齿,锯齿或多或少,或深或浅。藻体中下部叶片常分叉,近基部叶片常见 1—3个,少数4—7个叉,中部叶片偶见1个,而顶端叶片无分叉。次生分枝圆柱形,由初生分枝的叶腋生出。气囊球形、椭圆形或卵形,具圆柱形、亚圆柱形或叶状细柄,柄或长或短,顶端圆钝或具小刺状突起,偶见叶状,表面多毛窝,由叶腋生出。此时藻体已多见生殖托,但尚不易肉眼识别雌雄。生殖托圆柱形,顶端尖,表面光滑,疣状,由叶腋生出。

图1 潮间带自然栖息的雷州马尾藻Fig.1 S.leizhouense inhabitant in intertidal area
结果表明,藻体在同一固着器上的主干数为(6.00±2.96)个,同一主干的初生分枝数为(3.11±0.96)个,生殖托分枝(2.50±0.51)回(表1)。
2.1.2 生殖期 雷州马尾藻藻体可明显区分雌、雄个体(图3),雌、雄异体,生殖托呈复总状托序,具一短柄。雌生殖托(图4a)短,基部圆柱形,顶端多为二叉形,偶见三叉形,末端尖细,表面疣状,无刺,分枝1—3回。雄生殖托(图4b)呈长圆柱形,顶端无分枝,末端尖细,表面疣状,无刺,分枝3—4回。雌、雄体出现生殖托的位置距初生分枝基部的距离分别为(11.81±4.85)cm和(17.24±10.88)cm,雌、雄体次生分枝上托枝数分别介于16—39个和17—35个之间,平均分别为27.95±6.21个与25.94±5.01个。
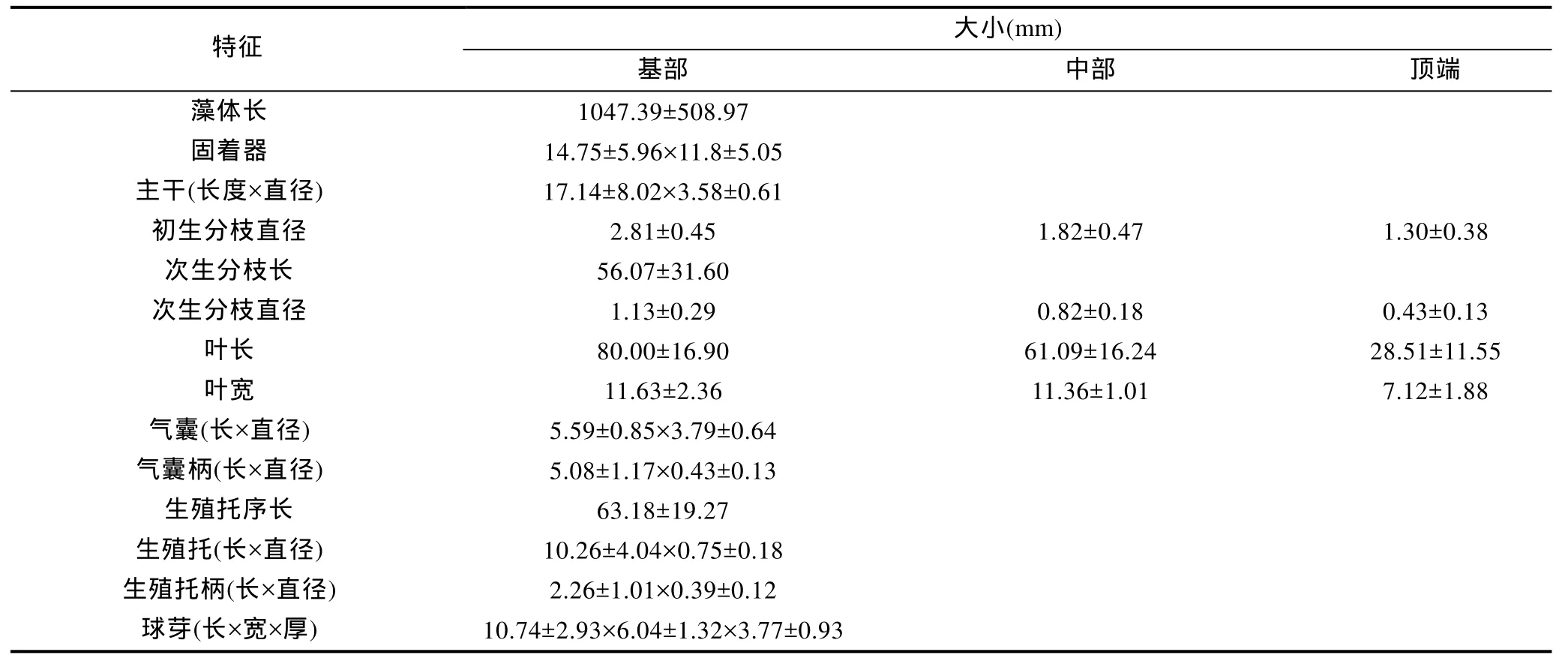
表1 雷州马尾藻的外部形态特征统计结果Tab.1 Statistical results of morphological observations to S.leizhouense

图2 雷州马尾藻各部位的形态观察Fig.2 Morphological observations of different parts of S.leizhouense

图3 雷州马尾藻的雌体(a)与雄体(b)Fig.3 Female(a)and male(b)individual of S.leizhouensis

图4 雷州马尾藻的雌生殖托(a)与雄生殖托(b)Fig.4 Female(a)and male(b)receptacles on S.leizhouensis
结果表明(表2),雷州马尾藻的雄性个体比雌性个体重。虽然雌体的初生分枝较宽,次生分枝较长、藻体中下部枝间距较小,球芽、藻叶与气囊皆较大,但由于其生殖托较小,成熟雄性生殖托长度平均为雌性的2.48倍,托柄长平均为雌性的2.24倍,单棵雄性藻体比雌性藻体重18.7%。

表2 雷州马尾藻藻体的外部形态特征统计结果Tab.2 Statistics of morphological features of male and female thalli of S.leizhouense
2.2 内部结构观察
2.2.1 营养体切面观察 雷州马尾藻分枝的横切面结构由表皮层、皮层和髓部三部分组成(图5)。表皮层由 1—3层含色素的细胞组成,细胞长多角形,大小为(13.56±4.01)μm×(10.54±2.39)μm;皮层细胞排列不甚规则,由 10.80±1.99层圆形、椭圆形或多角形细胞组成,大小不一,甚至相差悬殊,平均为(37.50±16.74)μm×(28.68±11.33)μm;髓部细胞比表皮细胞略大,切面观近圆形,大小为(17.23±5.06)μm×(14.31±3.98)μm。 藻 体 外 被 胶 质 层 ,厚(9.55±3.36)μm。

图5 雷州马尾藻分枝的横切面观Fig.5 The transverse section of S.leizhouense branches
2.2.2 生殖托切面观察 雌、雄生殖托的横切面结构中,表皮细胞都是由一层近方形细胞组成,雄托早期表皮细胞比雌性略小,而后期略大;皮层部位形成许多生殖窝,雌窝比雄窝大,窝外皮层细胞较营养期少,而窝内多见滋养丝细胞,发育早期雄性的滋养丝细胞较雌性的约大17%,而中、后期分别是雌性的 57.6%和 57.2%;髓部面积较营养期小,细胞数目少(表3,图6)。雌性生殖托的胶质层比雄性的厚18.7%。

表3 雷州马尾藻生殖托的横切面结构特征统计结果Tab.3 Statistics of transverse section of S.leizhouense receptacles

图6 雷州马尾藻雌、雄生殖托的横切面观Fig.6 The transverse section of female and male receptacles in S.leizhouense
2.3 雷州马尾藻的分类地位讨论
通过雷州马尾藻形态结构、生殖托(有一短柄,柄部长出小藻叶或小气囊)的观察,以及相关文献对比分析(曾呈奎等,2000),我们建议雷州马尾藻属于马尾藻属、叶托混生组(SectionZygocarpicaeSetchell)、拟叶托混生亚组(SubsectionPseudozygocapicaeSetchell)。
3 讨论
3.1 雷州马尾藻的形态结构
曾呈奎等(2000)在《中国海藻志》中对我国马尾藻属的分类依据是藻体的外部形态,主要包括固着器形状,主干长度、分叉与否、表面光滑与否,分枝数目、形状、长度、间隔,叶的形状、是否分叉、长度、宽度、中肋、毛窝与边缘形状,气囊的形状、大小、顶端形状与表面毛窝多少,生殖托的形状、长度、表面光滑与否,生殖托与叶、气囊着生方式,雌雄同体或异体等来判断,但并未涉及藻体内部结构,并且所有特征描述均缺乏统计分析。Mattio等(2008)研究了法国产的马尾藻形态并进行统计分析,但仅用叶子(长度×宽度)、气囊(直径)和生殖托(长度)3个参数的平均值。Wong等(2004)也采用统计学方法对棒托马尾藻(S.baccularia)与匍枝马尾藻(S.polycystum)的部分形态特征进行了分析。虽然Yoshida等(1983)对日本产的 1种马尾藻,以及 Noiraksar等(2008)对泰国产的10种马尾藻的生殖托进行了横切面观察,但仅有图片,未有统计结果。由此可见,马尾藻的形态结构研究既有定性描述也有定量研究,相关结果虽可相互参考,但相关的形态分类标准还需要讨论与统一。
3.2 雷州马尾藻分类地位的讨论
曾呈奎等(2000)将我国马尾藻属划分为3个亚属,其中叶枝亚属有4个种,反曲叶亚属16个种,真马尾藻亚属110个种,其后该方面未见新种报道。根据雷州马尾藻茎、藻叶有无明显分化,茎扁压、亚圆柱形和圆柱形,藻叶扁平,基部分枝或藻叶一般不向下反曲,从生殖托的基部长出小藻叶或小气囊等特征,我们认为雷州马尾藻属于真马尾藻亚属、叶托混生组、拟叶托混生亚组。
雷州马尾藻雌、雄异株,生殖托表面光滑,藻叶较厚,表面具有毛窝,应与瓦氏马尾藻(S.vachellianum)接近,但其形态特征又与瓦氏马尾藻的描述有较大出入,主要依据是:1.曾呈奎等(2000)未明确后者的初生分枝数,一般认为一个主干上仅有一个初生分枝;王伟定(2003)描述“主枝2—3条”,而雷州马尾藻同一主干上辐射状分布有2—4个初生分枝;2.瓦氏马尾藻无球芽,而雷州马尾藻有;3.瓦氏马尾藻中、下部藻叶无分叉,而雷州马尾藻有;4.瓦氏马尾藻生殖托为典型的圆锥花序,且雌、雄生殖托外形相似,而雷州马尾藻为总状托序,并且雌、雄生殖托除长短差异外,外形也明显不同。
通过对比分类地位相同的相关种类,我们发现雷州马尾藻与鳞茎马尾藻(S.bulbiferum)外形比较相似,二者同属于真马尾藻亚属、叶托混生组、拟叶托混生亚组,鳞茎马尾藻的特征描述与本种也极相近,但雷州马尾藻属于雌、雄异体,而鳞茎马尾藻属于雌、雄同体。
曾呈奎等(2000)将雷州马尾藻按生殖托序内没有小气囊或小藻叶归为滑托组(SectionMalacocarpicae),根据其生殖托总状排列,托柄无生殖窝,小枝表面光滑,生殖托较长,表面多缢缩,归为总状托序亚组(Racemosae),荚托系(SeriesSiliquosae)。滑托组、荚托系的亨氏马尾藻(S.henslowianum)在中国南海分布较广(曾呈奎等,2000),并且相关研究较多(李春莲,20111);Zouet al,2011;Weiet al,2012)。我们通过对比发现,雷州马尾藻与亨氏马尾藻的形态差异较大,主要表现为:1.雷州马尾藻有球芽,而亨氏马尾藻无;2.雷州马尾藻的初生分枝中、下部扁压或扁平,而亨氏马尾藻的呈圆柱状;3.雷州马尾藻基部的藻叶多呈分裂状,而亨氏马尾藻的不分裂;4.雷州马尾藻的生殖托顶端常分叉,而亨氏马尾藻的多单生。因此我们认为曾呈奎等(2000)将雷州马尾藻归为滑托组欠准确,因为本种生殖托的基部明显有小气囊或小藻叶,建议修改、完善该种的分类地位。
通过统计分析我们也发现,《中国海藻志》中对雷州马尾藻形态特征的描述欠妥。主要体现在藻体及各组成部分的大小方面。因为本研究观测到的数据明显比前期记载的大,所采集的雷州马尾藻个体长达3m以上,而记载“雷州马尾藻藻体高至50cm”(曾呈奎等,2000);两者从主干上生出初生分枝和一些芽,所记载的“芽较短,高约1cm,直径为3—5mm”。本研究中雷州马尾藻的芽长达1.4cm,比记载的高40%,直径达7.2mm,约是记载的2倍;其他特征如藻叶、气囊和生殖托大小等的描述也有出入,书中记载的雷州马尾藻初生分枝圆柱状,藻叶较小,藻叶较薄等特征与研究的雷州马尾藻形态特征明显不符,本种的初生分枝中、下部皆扁压或扁平,顶部呈圆柱状;基部藻叶长可达16.3cm,宽可达1.9cm,且明显较厚。综上所述,我们认为曾呈奎等(2000)所述的雷州马尾藻标本特征与采集样本的数量及地点有关,这可能与当时海藻样本采集量少有关,缺乏统计结果。另外,曾呈奎等(2000)所采集的标本是二十世纪七、八十年代的,经过近三、四十年潮间带各种条件的变化,雷州马尾藻的形态等各方面也可能发生较大的变化。曾呈奎等(2000)描述的标本采自广东硇洲岛海域,而本种采自徐闻海域,两地海面相距约86海里,两地的同种马尾藻是否存在地理差异,还需进一步研究。
4 结论
4.1 雷州马尾藻形态特征描述
藻体呈黄褐色至褐色,由固着器、主干、初生分枝、球芽、叶片、次生分枝、气囊和生殖托组成,单生或丛生,长达3m以上。固着器盘状,大小为(14.75±5.96)mm×(11.8±5.05)mm。主干圆柱形,表面瘤状,长(17.14±8.02)mm,直径(3.58±0.61)mm。初生分枝2—4条,由主干顶部长出,呈辐射状排列,下部(约占藻体长度的2/3)扁压或扁平,直径(1.82±0.47)—(2.81±0.45)mm,上部(约占藻体长度的1/3)圆柱形或亚圆柱形,表面光滑,直径(1.30±0.38)mm。初生分枝基部分布有2—5个椭圆形球芽,表面光滑,长(10.74±2.93)mm,宽(6.04± 1.32)mm,厚(3.77±0.93)mm。叶片长披针形,具一短柄,基部略楔形,顶 端 圆 钝 或 尖 细 ,叶 长(28.51±11.55)—(80.00±16.90)mm;叶宽(7.12± 1.88)—(11.63±2.36)mm。叶上中肋明显,并延伸至叶尖,中肋两侧多毛窝,叶片边缘全缘或有锯齿,锯齿或多或少,或深或浅。藻体基部叶片常分叉,近基部叶片常见1—3个,少数4—7个,中部及顶端叶片偶见1个分叉。次生分枝圆柱形,由初生分枝的叶腋生出,长(56.07±31.60)mm,直径(0.43± 0.13)—(1.13±0.29)mm。气囊球形、椭圆形或卵形,具圆柱形、亚圆柱形或叶状细柄,柄或长或短,顶端圆钝或具小刺状突起,偶见叶状,表面多毛窝,由 叶 腋 生出 ,长(5.59±0.85)—(7.18± 1.03)mm,直径(3.79±0.64)—(5.30±0.59)mm,柄 长(5.08±1.17)—(5.30±1.86)mm,柄径(0.43±0.13)—(0.80±0.44)mm。雌雄异体,生殖托呈复总状托序,具一短柄。雌生殖托短,基部圆柱形,顶端多为二叉形,偶见三叉形,末端尖细,表面疣状,无刺,分枝1—3回,长(7.96±1.84)mm,宽(0.99±0.11)mm,托 柄 长(1.07±0.43)mm,宽(0.42±0.09)mm。雄生殖托长圆柱形,通常顶端无分枝,末端尖细,表面疣状,无刺,分枝3—4回,长(19.78±5.80)mm,宽(1.00±0.17)mm,托柄长(2.40±1.03)mm,宽(0.41±0.09)mm。雌、雄体出现生殖托的位置距初生分枝基部的距离分别为(11.81±4.85)cm和(17.24±10.88)cm,雌、雄体次生分枝上托枝数分别介于16—39个和17—35之间,平均分别为(27.95±6.21)个与(25.94±5.01)个。多生长在低潮带及潮下带1—2m深处。
4.2 雷州马尾藻及相近种检索表
1.藻叶扁平,气囊从叶腋中长出(马尾藻属Sargassum)···································································2
2.基部分枝或藻叶一般不向下反曲(真马尾藻亚属Sargassum)···························································3
3.从生殖托的基部长出小藻叶或小气囊(拟叶托混生组Pseudozygocapicae)·····································4
3.生殖托序内没有小藻叶或小气囊,生殖托表面光滑(滑托组Pseudozygocapicae)····························7
4.藻体雌雄同株,基部有鳞茎··········································································鳞茎马尾藻S.bulbiferum
4.藻体雌雄异株·················································5
5.生殖托表面光滑·············································6
6.藻叶较厚,基部藻叶不分裂,初生分枝基部无球芽·······························瓦氏马尾藻S.vachellianum
6.藻叶较厚,基部藻叶常常分裂,初生分枝基部有球芽······························雷州马尾藻S.leizhouense
7.生殖托总状排列、较长,初生分枝圆柱状,藻叶较小、不分裂················亨氏马尾藻S.henslowianum
王伟定,2003.浙江省马尾藻属和羊栖菜属的调查研究.上海水产大学学报,12(3):227—232
王 博,黄 和,熊皓平等,2010.硇洲马尾藻的营养成分分析与评价.食品研究与开发,31(10):195—197
刘东超,赵素芬,2006a.马尾藻—具有人工栽培前景的大型经济海藻.海洋与渔业,11:18—20
刘东超,赵素芬,谢青明,2006b.马尾藻池塘人工养殖技术初步研究.中国水产,1:63—64
李来好,杨贤庆,吴燕燕等,1997.马尾藻的营养成分分析和营养学评价.青岛海洋大学学报,27(3):319—325
李春莲,2011.低分子量亨氏马尾藻岩藻聚糖硫酸酯的制备及抗肿瘤活性研究.湛江:广东海洋大学硕士论文,1—74
赵素芬,孙会强,袁振江等,2006.硇洲岛春季底栖海藻资源.上海水产大学学报,15(3):321—327
赵素芬,罗世菊,孙会强等,2012.海藻与海藻栽培学.北京:国防工业出版社,178—222
曾呈奎,张俊甫,1959.关于几种褐藻在中国沿岸的不连续分布.海洋与湖沼,2(2):86—92
曾呈奎,陆保仁,2000.中国海藻志.第三卷 褐藻门,第二册墨角藻目.北京:科学出版社,12—124
Agardh C A,1820.Species Algarum Rite Cognitae,Cum Synonymis,Differentiis Specificis et Descriptionibus Succinctis.vol.1.Berling,Lund:168
Agardh J G,1848.Species Genera et Ordines Algarum,Seu Descriptiones Succinctae Specierum,Generum et Ordinum,Quibus Algarum Regnum Constituitur.Algas Fucoideas Complectens,vol.1.Gleerup,C W K,Lund:363
Agardh J G,1889.Species Sargassorum Australiae descriptae et dispositae.Öfversigt af Kongl.Svenska Vetenskaps-Adademiens Handlingar,23:1—133
Mattio L,Payri C E,2011.190 years ofSargassumtaxonomy,facing the advent of DNA phylogenies.The Botanical Review,77(1):31—70
Mattio L,Payri C E,Stiger-Pouvreau V,2008.Taxonomic revision ofSargassum(Fucales,Phaeophyceae)from French Polynesia based on morphological and molecular analyses.Journal of Phycology,44(6):1541—1555
Noiraksar T,Ajisaka T,2008.Taxonomy and distribution ofSargassum(Phaeophyceae)in the Gulf o f Thailang.Journal of Applied Phycology,20(5):513—527
Sun J Z,Ning X R,Le F Fet al,2010.Long term changes of biodiversity of benthic macroalgae in the intertidal zone of the Nanji islands.Acta Ecologica Sinica.30(2):106—112
Tseng C K,1985.SargassumsectionPhyllocystaesect.nov.,a new section ofSargassumsubgenusBatrophycus.In:Abbot I A,Norris J N eds.Taxonomy of Economic Seaweed:With Reference to Some Pacific and Caribbean Species.vol.1.California Sea Grant,La Jolla,CA:15
Wei M Y,Li S D,Chen J Xet al,2012.Chemical constituents of the brown algaSargassum henslowianumcollected from the south China Sea.Chemistry of Natural Compounds,48(4):677—678
Wong C L,Gan S Y,Phang S M,2004.Morphological and molecular characterisation and differentiation ofSargassum bacculariaandS.polycystum(Phaeophyta).Journal of Applied Phycology,16(6):439—445
Womersley H B S,1954.Australian species ofSargassum,subgenusPhyllotrichia.Australian Journal of Botany,2(3):337—354
Xie E Y,Liu D C,Liu Cet al,2013.Artificial seed production and cultivation of the edible brown algaSargassum naozhouenseTseng et Lu.Journal of Applied Phycology,25(2):513—522
Ye H,Zhou C H,Sun Yet al,2009.Antioxidant activities in vitro of ethanol extract from brown seaweedSargassum pallidum.Europe Food Research Technology,230(1):101—109
Yoshida T,Konno T,1983.Taxonomic study onSargassum sagamianumYendo and related species(Phaeophyta,Fucales).The Botanical Magazine,96(3):145—157
Zou D H,Gao K S,Chen W Z,2011.Photosynthetic carbon acquisition inSargassumhenslowianum(Fucales,Phaeophyta),with special reference to the comparison between the vegetative and reproductive tissues.Photosynthesis research,107(2):159—168
猜你喜欢
中国海洋大学学报(自然科学版)(2022年8期)2022-08-15 06:35:20
浙江农业学报(2020年1期)2020-03-07 05:07:12
大自然探索(2019年9期)2019-12-13 07:25:02
儿童故事画报·自然探秘(2017年10期)2018-03-15 17:38:02
浙江农业科学(2017年8期)2017-08-30 12:48:08
科技创新与应用(2017年20期)2017-07-15 20:43:02
浙江农业科学(2016年5期)2016-12-07 07:03:27
广东海洋大学学报(2015年4期)2016-01-13 08:39:29
——系统分类与检索*
寄生虫与医学昆虫学报(2015年3期)2015-11-12 00:32:38
湖泊科学(2015年4期)2015-02-17 06:43:49
