
不同种源卷丹染色体数目变异1)
2014-03-08 05:44杨利平
东北林业大学学报 2014年11期
唐 彪 杨利平 侯 菲
(长江师范学院,重庆,408100)
责任编辑:任 俐。
卷丹(Lilium lancifolium Thunb.)分布极广,自俄罗斯的西伯利亚到我国的西北、东北、华北、华中、华东和西南等地,日本、朝鲜也有分布[1]。研究认为,卷丹在日本、朝鲜半岛和中国东北的自然界有天然多倍体存在[2-7]。Noda[6]认为,日本多倍体卷丹是由二倍体卷丹和大花卷丹(L.maximowiczii)形成的异源三倍体,而分布在韩国的多倍体卷丹有同源三倍体居群存在[7]。在韩国的卷丹居群调查表明,二倍体集中分布西部东南方向沿岸和岛屿的悬崖峭壁,三倍体则集中分布于内陆地区[7],而日本对马岛的二倍体和三倍体共同存在,且二倍体珠芽形态与三倍体极为相似[5]。Hwang[8]利用FISH 技术对二倍体与三倍体卷丹染色体组进行精准定位,明确二者之间的染色体对应关系。中国产卷丹几乎都是三倍体[9-13],陕西汉中种源地卷丹为同源三倍体[14]。但中国植物志中有关卷丹有果实的描述,又给研究者对中国产卷丹是否存在二倍体或其他情况提供更多的思考[1]。卷丹在我国广为栽培,不仅是我国百合药材的主要来源,也是三大食用百合之一,同时作为球根花卉在园林中普遍应用[15-17]。有证据显示,经过长期的栽培和民间选育,卷丹已经出现遗传分化,如宜兴百合和南京紫金山百合都是卷丹的栽培变种,但其染色体变化未见报道[18-19]。目前,对广泛分布卷丹染色体一些有限的研究报道,还远不能全面揭示该物种染色体数目变异全部信息和在我国的分布状态,因此,对不同种源产地卷丹种质染色体倍性研究分析,可为该物种有效保护和育种应用提供更加丰富的遗传背景资料。
1 材料与方法
试验所用卷丹分别来自江苏省宜兴(JD -Y)、重庆市武隆(JD-W)、黑龙江省齐齐哈尔(JD-Q)、黑龙江省哈尔滨(JD-H)和江西省庐山(JD-L)。
采用植物染色体常规压片法观察染色体数目[20]。将无菌苗25 ℃恒温生根培养,至根长1 cm左右,切取根尖放入0.2%秋水仙素溶液中,预处理24 h,卡诺固定液(V(95%乙醇)∶ V(冰醋酸)=3∶1)固定6 h,1 mol·L-1盐酸60 ℃恒温水浴解离6 min,卡宝品红染色8 min 后压片。显微镜下镜检,每种源统计30 个以上清晰分裂相的细胞染色体数目,并拍照。
2 结果与分析
宜兴种源卷丹的染色体数目变异范围为28~40,其中36 条染色体细胞数占37.21%,37 条染色体细胞数占18.60%,35 条染色体细胞数占13.95%,其他染色体数目所占比例较小;重庆市武隆种源卷丹的染色体数目变异范围为27~39,其中36条染色体细胞数占37.10%,35 条染色体细胞数占22.58%,34 和38 条染色体细胞数所占比例接近10%;黑龙江省齐齐哈尔种源卷丹的染色体数目变异范围为32~39,其中36 条染色体细胞数占39.58%,34、35 和37 条染色体细胞数所占比例都超过10%;黑龙江省哈尔滨种源卷丹的染色体数目变异范围为28~36,其中36 条染色体细胞数占54.76%,34 条染色体细胞数所占比例接近20%,其他染色体数目的比例较小;江西省庐山种源卷丹的染色体数目变异范围为24~38,其中36 条染色体细胞数占32.81%,35 条染色体细胞数所占比例为26.56%,34、37 和38 条染色体细胞数所占比例接近或超过10%,其他染色体数目的比例较小,包括1.56%的二倍体细胞(2n=24)(表1,图1)。

表1 卷丹体细胞染色体数目及所占比例
按着李懋学等[21]的标准,85%以上的细胞具恒定一致的染色体数,即可认为是该植物的染色体数目。如果观察材料系混倍体,则应如实记录其染色体数的变异范围和各类细胞的数量或百分比。在对卷丹的根尖细胞染色体数目统计分析发现,5 个种源中,2n=3x =36 细胞比例虽然占有绝对的多数,但都没有超过85%,个体染色体数目成单个或几个的增添或减少,因此,所观察的5 个种源的卷丹都不能称为三倍体或整倍体,应该为整倍与非整倍的混倍体。
3 结束语
通常在植物界的某些属甚至某些科往往具有同一染色体基数,而在种内染色体数目具有相对的稳定性。以往的研究中已经确认百合属的染色体基数为n=12,多数种类2n=2x=24,只有卷丹有二倍体(2n=2x=24)和三倍体(2n =3x =36)两种染色体数目形态[6-7,22]。近年有研究者观察到,东北产3个居群卷丹中同时存在非整倍性变异和整倍性变异,观察到有单倍体细胞,但36 条染色体细胞占89%,该居群仍为三倍体[12]。研究者在对中国产百合52 个居群的30 个分类群调查时发现,所有居群染色体基数为12,为二倍体,卷丹为三倍体,首次发现有一个居群的淡黄花百合(L.sulphureum)为非整倍体(2n=23)[13]。
笔者从事野外考察工作中,遇到的卷丹三倍体几乎不是在路旁,就是在寺庙旁,而且其中个体数量不大[11],这与Kim et al.[7]同源三倍体集中分布于内陆地区的路旁、田头和丘陵荒地的结论基本相符。在东北林业大学、中国科学院沈阳应用生态研究所、中国科学院北京植物研究所和东北师范大学植物标本馆查阅百合属标本时也发现,该属其他种类生长在野生环境,只有卷丹的生境为村庄旁、小路旁或寺庙旁等与人类活动及其相关的地点。
经过对卷丹野外调查、栽培地实地考察和标本馆查阅同时发现,卷丹的形态特征变化极大,包括株高、茎的颜色(紫色或绿色)、茎上有无棱或毛、节间长短、叶形(长披针形或阔披针形)、珠芽大小及形态等。目前,卷丹的形态分化、遗传分化及形态与染色体的相关性等研究还未见报道,但可以初步推断产生卷丹多样性变异的几种可能:以卷丹为亲本衍生出的品种很多[23-24],人类活动将卷丹的不同品种从不同途径带到我国各地,由于卷丹及其品种的适应性很强,有些变成逸出种(半野生种),慢慢形成了现在的卷丹如此多样化的局面;或是卷丹长期生长在异质生境中,形成了遗传和形态较为丰富的生态类型;或长期的栽培和民间选育出现的遗传分化[18]。
三倍体卷丹不结实,自然条件下依靠地下鳞茎周围生出的小鳞茎或地上茎上部叶腋处的珠芽来进行无性繁殖,因此,混倍体群体或个体可以无性方式延续种群。染色体结构变异为百合属进化的一条主要途径,多倍化是其进化的一条支线[12],而非整倍 化也可以成为百合属中的一种进化途径。
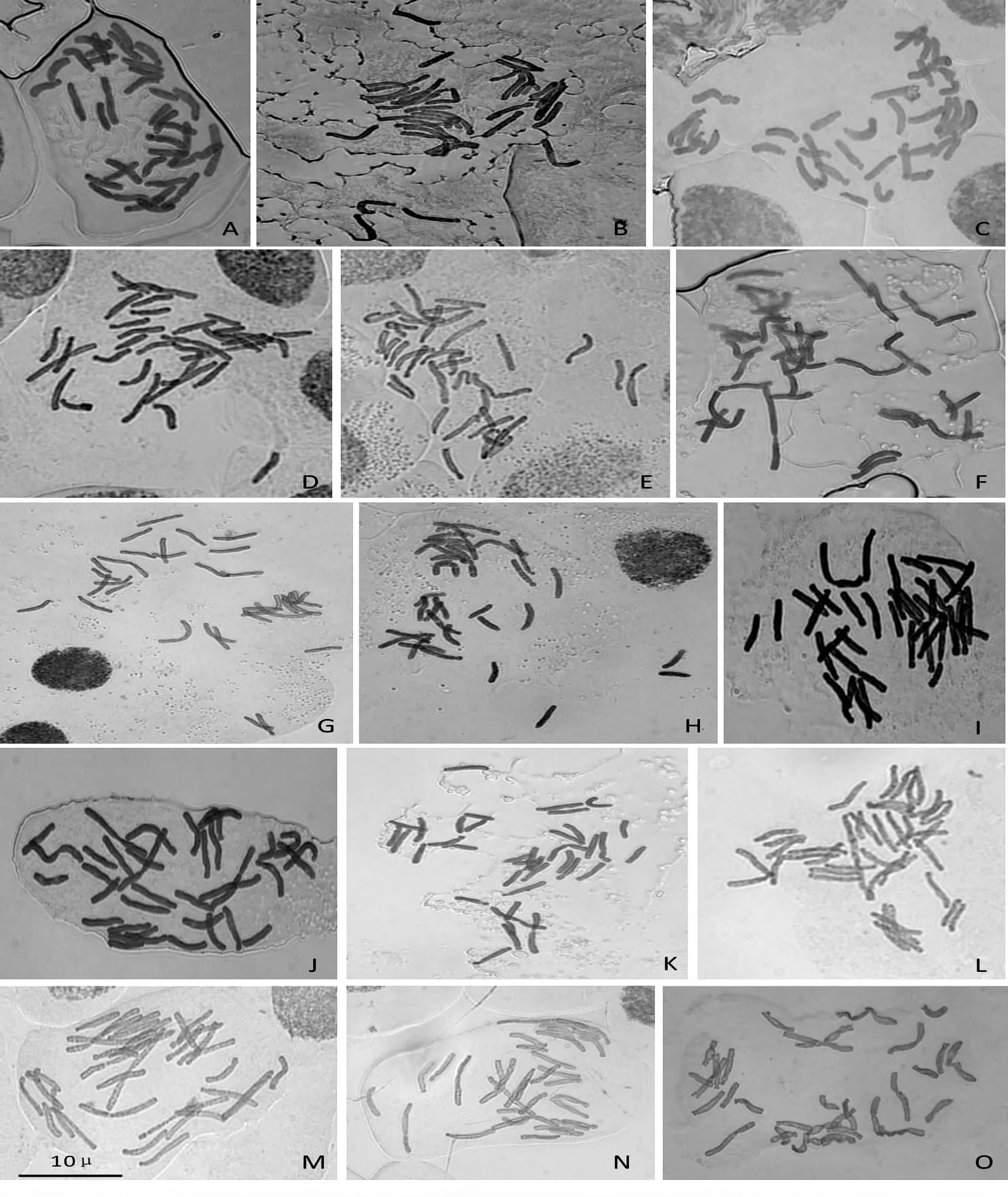
图1 卷丹根尖染色体数目
[1] 汪发缵,唐进.中国植物志:第14 卷:百合科[M].北京:科学出版社,1980:159 -165.
[2] Jonathan J W.Cytological Studies of Lilium tigrinum[J].Botanical Gazette,1940,101(3):550 -581.
[3] Stewart R N,Baraford R.The nature of polyploidy in Lilium tigrinum[J].American Journal of Botany,1943,30(1):783 -794.
[4] Peter B M.Synaptinemal complexes of Lilium tigrinum (triploid)sporocytes[J].Canadian Journal of Genetics and Cytology,1968,10(4):799 -807.
[5] Noda S.Choromosomes of diploid and triploid forms found in the natural populations of Tiger Lily in Tsushima[J].Bot Mag Tokyo,1978,91:279 -283.
[6] Noda S.Cytogenetic behavior,chromosomal differentiations and geographic distribution in Lilium lancifolium (Liliaceae)[J].Plant Species Biology,1986,1(1):69 -78.
[7] Kim J H,Kyung H Y,Choi Y S,et al.Geographic distribution and habitat differentiation in diploid and triploid Lilium lancifolium of South Korea[J].Fac Agr Kyushu Univ,2006,51(2):239-243.
[8] Hwang Y J,Kim H H,Kim J B,et al.Karyotype analysis of Lilium tigrinum by FISH[J].Horticulture,Environment and Biotechnology,2011,52(3):292 -297.
[9] 仲裕泉,李浩兵,王克娟.南京紫金山百合的核型和C -带带型分析[J].江苏农业学报,1993,9(3):8 -12.
[10] 图力古尔,刘立波.吉林省产5 种百合的核型研究[J].武汉植物学研究,1996,14(1):6 -12.
[11] 杨利平,丁冰,刘香环,等.东北百合属植物的细胞遗传多样性[J].东北林业大学学报,1996,24(5):19 -23.
[12] 荣立苹,雷家军,毕晓颖,等.8 种野生百合染色体数目的变异[J].东北林业大学学报,2009,37(9):48 -50.
[13] Gao Yundong,Zhou Songdong,He Xingjin.Karyotype studies in thirty-two species of Lilium (Liliaceae)from China[J].Nordic Journal of Botany,2011,29(6):746 -761.
[14] 刘晓丽,贾桂霞.卷丹小孢子母细胞减数分裂染色体行为的研究[J].植物遗传资源学报,2013,14(1):151 -157.
[15] 国家药典委员会编.中华人民共和国药典:一部[M].北京:中国医药科学技术出版社,2010:123.
[16] 柴春燕,胡东旭.宜兴百合特性及高产栽培技术[J].广西园艺,2003(3):24 -25.
[17] 车景超,丰杰,辛宁.卷丹和百合化学成分与提取方法研究进展[J].时珍国医国药,2008,19(6):1500 -1501.
[18] 刘光欣,胡凤荣,席梦利,等.卷丹与宜兴百合的C -带比较[J].南京晓庄学院学报,2009(6):63 -65.
[19] 仲裕泉,李浩兵,王克娟.南京紫金山百合的核型和C -带带型分析[J].江苏农业学报,1993,9(3):8 -12.
[20] 李懋学,张斅方.植物染色体研究技术[M].哈尔滨:东北林业大学出版社,1991:31 -58.
[21] 李懋学,陈瑞阳.关于植物核型的标准化问题[J].武汉植物研究,1985,3(4):297 -302.
[22] 洪德元.植物细胞分类学[M].北京:科学出版社,1990,91 -96.
[23] Whipker B E,McCall I,Krug B,et al.Flurprimidol preplant bulb soaks control growth of potted Lilium[J].Acta Hort,2011,900:79 -88.
[24] Barnes J,Whipker B,Buhler W,et al.Flurprimidol pre-plant bulb soaks control growth of Lilium lancifolium‘Orange Tiger’[R].Asheville,North Carolina,USA:Proceedings of the 36th Annual meeting of the Plant Growth Regulation Society of America,2009.
猜你喜欢
中国海洋大学学报(自然科学版)(2023年2期)2023-02-21
河北科技师范学院学报(2022年2期)2022-08-26
科学养鱼(2021年11期)2022-01-11
浙江中医药大学学报(2021年6期)2021-07-12
中国粮油学报(2020年12期)2021-01-09
热带农业科技(2020年3期)2020-12-15
食品与生活(2019年12期)2019-05-15
江苏农业科学(2017年16期)2017-10-27
现代农业科技(2017年8期)2017-06-10
中国果业信息(2015年6期)2015-01-23
