
新疆伊犁河岸小叶白蜡复叶构件的表型可塑性1)
2014-03-08 05:44努尔买买提杨晓绒贾娜尔
东北林业大学学报 2014年11期
努尔买买提 张 维 杨晓绒 贾娜尔
(伊犁师范学院,伊宁,835000)
责任编辑:任 俐。
小叶白蜡(Fraxinus sogdiana)又名天山梣,属木樨科(Oleaceae)白蜡属植物,是珍贵的第三纪温带阔叶林残遗树种[1],其抗寒能力强,材质坚韧而有弹性,树形美观,它还是放养白蜡虫的优良树种。在我国成片分布的小叶白蜡原始林,仅见于新疆天山西部伊犁谷地,主要生长在喀什河下游河谷及河岸滩地。
表型可塑性是生物适应环境的重要方式[2-3],生物个体基因型通过产生多种表现型以适应不同的环境条件[4-5],表型可塑性在种群水平具有调节生长与物质分配策略的功能和意义[6]。叶片是植物对环境条件变化较为敏感的营养器官,具有物质生产功能[7]。植物生长发育的可塑性在时间上随不同的基因型、种群或物种而发生改变[8-9]。生长节律的变化是植物本身生理机能的体现,同时也受外界环境的影响。采用遗传上较为稳定、不易受环境影响的性状,如叶、花等性状研究表型可塑性,可以揭示群体的变异大小和遗传规律,评价其遗传多样性水平,进而为群体的保护提供理论支持[10-11]。
表型可塑性使生物体在多变的环境中生存,增加了生物体的生态幅,避免个体死亡或缓冲分布范围的变化[2,12],因此,众多学者从生态学角度对表型可塑性进行了探讨[5,9-10,12-13]。前人对小叶白蜡的生物学特性、光合特征、抗逆性、病害防治、种子成分及发芽率、育苗及移植栽培技术[14-20]等已有研究,其复叶的表型变异性尚未见报道。文中以新疆小叶白蜡自然保护区小叶白蜡的复叶为研究对象,通过大样本取样测量,分析5 种不同小叶数量复叶的表型可塑性,拟为植物生长调节、适应与进化等深入研究提供科学积累,为干旱区植物器官适应的可塑性提供素材,并为小叶白蜡植物资源的利用和保护提供参考。
1 研究区自然概况
小叶白蜡自然保护区位于新疆伊宁市东南部50 km,喀什河汇入伊犁河河口地带(81°51'~81°54'E,43°44'~43°48'N),地势平坦,平均海拔802 m,小叶白蜡集中分布面积约400 hm2。该区属北温带大陆性半干旱气候,年日照时间2 800 h,年平均气温8.2 ℃,1月份平均气温-12.8 ℃,7月份平均气温24.7 ℃,极端低温-39.2 ℃,极端高温37.5 ℃,≥10 ℃积温3 684 ℃,无霜期约160 d,年降水量480 mm,年蒸发量1 420 mm,相对湿度60%~70%,地下水位1.0~1.5 m,土壤类型为沙壤土或盐化潮土,pH 值8.0。小叶白蜡为保护区的优势树种,伴生乔木主要有密叶杨(Populus talassica)、榆树(Ulmus pumila)、多枝柽柳(Tamarix ramosissima)等;林下灌木主要有异果小檗(Berberis heteropoda)、铃铛刺(Halimodendron halodendron)等;草本植物主要有短柄草(Brachypodium sylvaticum)、草地早熟禾(Poa pratensis)、问荆(Equisetum arvense)、北艾(Artemisia vulgaris)等。
2 材料与方法
2012年8月中旬,在小叶白蜡自然保护区不同位置(东、南、西、北和保护区中央)各选择成年小叶白蜡树10 棵。在选定的目标树不同叶位随机采摘完整的成熟复叶,每种复叶各采摘30 个作为分析样本。小叶白蜡为奇数羽状复叶,常见有5 小叶、7 小叶、9 小叶、11 小叶和13 小叶5 种类型(单株小叶白蜡树主要生长一种或两种类型复叶,偶有三种类型复叶),用米尺测量各组复叶的叶轴长、复叶长、复叶宽,每片小叶的叶长和叶宽,单位为mm。
采用统计软件SPSS13.0 对测量数据分析处理和作图。不同复叶的相同构件用方差分析比较其差异性。研究表型可塑性的主要方法是分析反应范式[21],反应范式是指功能特征随环境梯度改变而改变的函数[21],对小叶白蜡的复叶长与叶轴长、复叶长与复叶宽、小叶长与小叶宽作散点图,并对复叶构件间的生长关系作回归分析。分析植物构件的异速生长或同速生长常采用线性函数、对数函数、幂函数和指数函数模拟[13,22-23],以这四种函数模拟小叶白蜡复叶的生长特征,并选用相关性最高的拟合方程作为其生长关系的定量描述模型,回归方程的拟合优度检验用判定系数R2实现,回归方程的显著性采用F 检验。作散点图和回归分析时,相同复叶的顶小叶和左、右小叶合并处理。
3 结果与分析
3.1 不同复叶构件的表型可塑性
经测量,小叶白蜡全体复叶样本的叶轴长47.9~203.7 mm,复叶长119.1~265.6 mm,复叶宽74.2~172.1 mm,小叶长13.6~95.9 mm,小叶宽10.2~48.1 mm。不同复叶构件的统计结果见表1。由表1可见,叶轴长:随小叶数增多而增大,13 小叶是5 小叶的2.3 倍;不同复叶间差异显著;复叶间的差异(变异系数)均大于复叶内的差异。复叶长:随小叶数增多而增大,13 小叶比5 小叶长72.0 mm;7 小叶和9 小叶差异不显著,其余均差异显著;复叶间的差异均大于复叶内的差异。复叶宽由大到小的顺序为7 小叶、9 小叶、5 小叶、11 小叶、13 小叶,除7 小叶外,其余复叶宽在复叶间的差异均小于复叶内的差异。小叶长以7 小叶最大,13 小叶最小;小叶宽以9 小叶最大,5 小叶最小。小叶长和小叶宽在复叶间的差异均小于复叶内的差异。
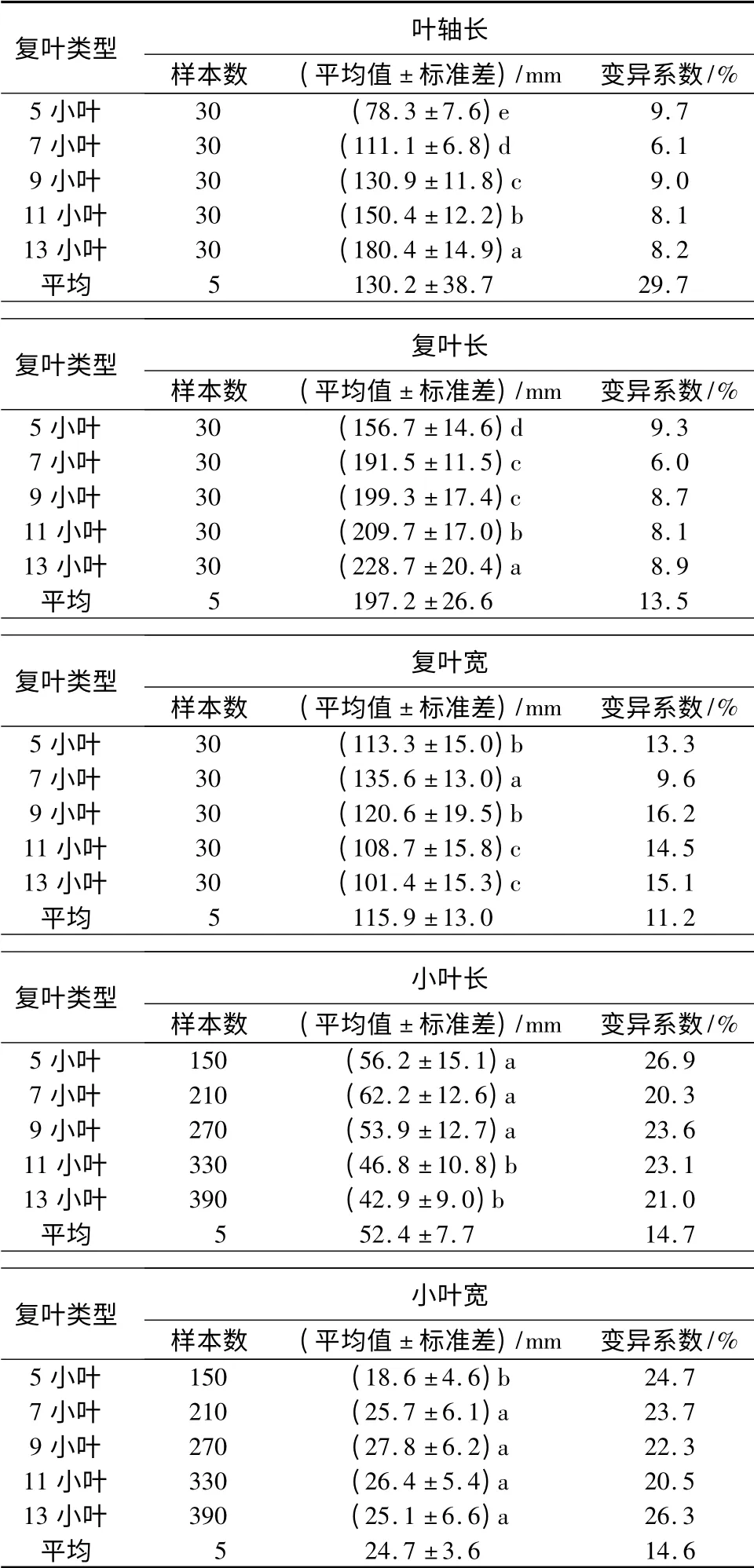
表1 小叶白蜡不同复叶构件的数量特征及其差异性
由标准差可见:叶轴长和复叶长以13 小叶离散程度最大,7 小叶最小;复叶宽以9 小叶最大,7 小叶最小;小叶长以5 小叶最大,13 小叶最小,而小叶宽以13 小叶最大,5 小叶最小。综上所述,小叶白蜡复叶构件在不同复叶间和复叶内均存在较大可塑性。
3.2 复叶长与叶轴长的生长模型
以叶轴长为横坐标,复叶长为纵坐标作散点图并赋拟合方程(图1)。小叶白蜡5 种复叶的叶轴长与复叶长均为异速生长型,其拟合方程均达到极显著(p <0.01)水平。其中5 小叶、7 小叶、9 小叶的复叶长与叶轴长的生长关系可用对数函数作为定量描述模型,由对数函数的性质可知,这3 种复叶的叶轴长在复叶生长发育的整个过程均增长较快,复叶长在生长的初期增长较快,随后增长逐渐变慢,最后趋于稳定;11 小叶、13 小叶的复叶长与叶轴长的生长关系可用指数函数作为定量描述模型,由指数函数的性质反映出这2 种复叶的叶轴长在复叶生长发育的整个过程均增长较慢,复叶长在生长初期增长较慢,随后逐渐加快生长。如果把5 种复叶看做一个样本,复叶长与叶轴长的生长关系可用指数函数作为定量描述模型。
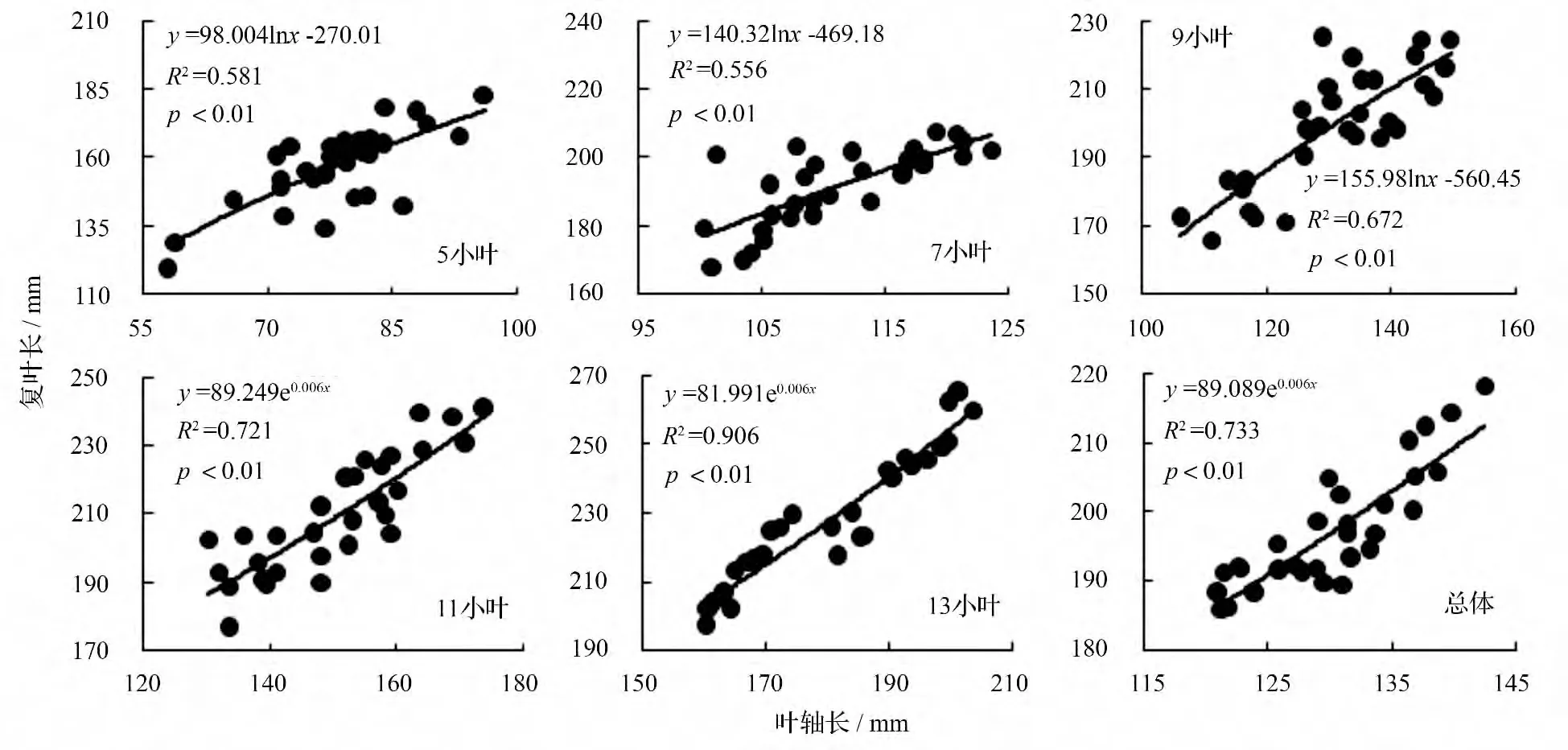
图1 复叶长与叶轴长的观测值及拟合曲线
3.3 复叶长与复叶宽的生长模型
以复叶宽为横坐标、复叶长为纵坐标作散点图并赋拟合方程(图2)。小叶白蜡5 种复叶的复叶长与复叶宽均为异速生长型,其拟合方程均达到极显著(p <0.01)水平。其中5、7 和9 小叶的复叶长与复叶宽的生长关系可用对数函数作为定量描述模型,即这3 种复叶的复叶宽在复叶生长发育的整个过程均增长较快,复叶长在生长的初期增长较快,随后增长逐渐变慢,最后趋于稳定;11 小叶、13 小叶的复叶长与复叶宽的生长关系可用指数函数描述,即这2 种复叶的复叶宽在复叶生长发育的整个过程均增长较慢,复叶长在生长初期增长较慢,随后逐渐加快增长。如果把5 种复叶看做一个样本,复叶长与复叶宽的生长关系可用指数函数作为定量描述模型。
3.4 小叶长与小叶宽的生长模型
以小叶宽为横坐标,小叶长为纵坐标作散点图并赋拟合方程(图3)。小叶白蜡5 种复叶的小叶长与小叶宽既有同速生长型,又有异速生长型,其拟合方程均达到显著(p <0.05)水平。其中5 小叶、9 小叶、13 小叶的小叶长与小叶宽的生长关系可用直线函数作为定量描述模型,即小叶长和小叶宽在复叶的整个生长发育过程中成比例生长;7、11 小叶的小叶长与小叶宽的生长关系可用幂函数作为描述模型,即这2 种复叶的小叶宽在复叶形成初期增长很快,小叶长增长较慢,过曲线拐点后,小叶宽增长非常缓慢,小叶长则快速增长。同样,如果把5 种复叶看做一个样本,小叶长与小叶宽的生长关系可用直线函数作为定量描述模型。
4 结论与讨论
表型可塑性往往具有适应和进化上的意义,如植物叶柄长度不仅影响树汁的传导,而且影响叶片的空间分布,从而对光的截留具有重要作用[24]。从平均值反映出,小叶白蜡不同复叶的叶轴长最大值是最小值的2.3 倍,复叶长和小叶宽最大值与最小值相差1.5 倍,复叶宽和小叶长最大值与最小值相差约1.4 倍;不同复叶构件有不同的离散程度(标准差),一部分构件在小叶数多的复叶离散程度大,另一部分构件在小叶数少的复叶离散程度大;叶轴长、复叶长在复叶间的差异(变异系数)均大于复叶内的差异,小叶长和小叶宽在复叶间的差异均小于复叶内的差异。以上三个方面的数量特征,揭示了小叶白蜡不同复叶有较强的可塑性。叶轴长随复叶的小叶数增多而增大,与叶轴具有养分疏导及支撑叶片扩张生长空间等多重功能相适应,这是小叶白蜡复叶为了适应其生理功能在生长发育上的一种权衡。
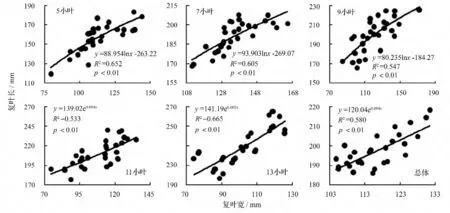
图2 复叶长与复叶宽的观测值及拟合曲线
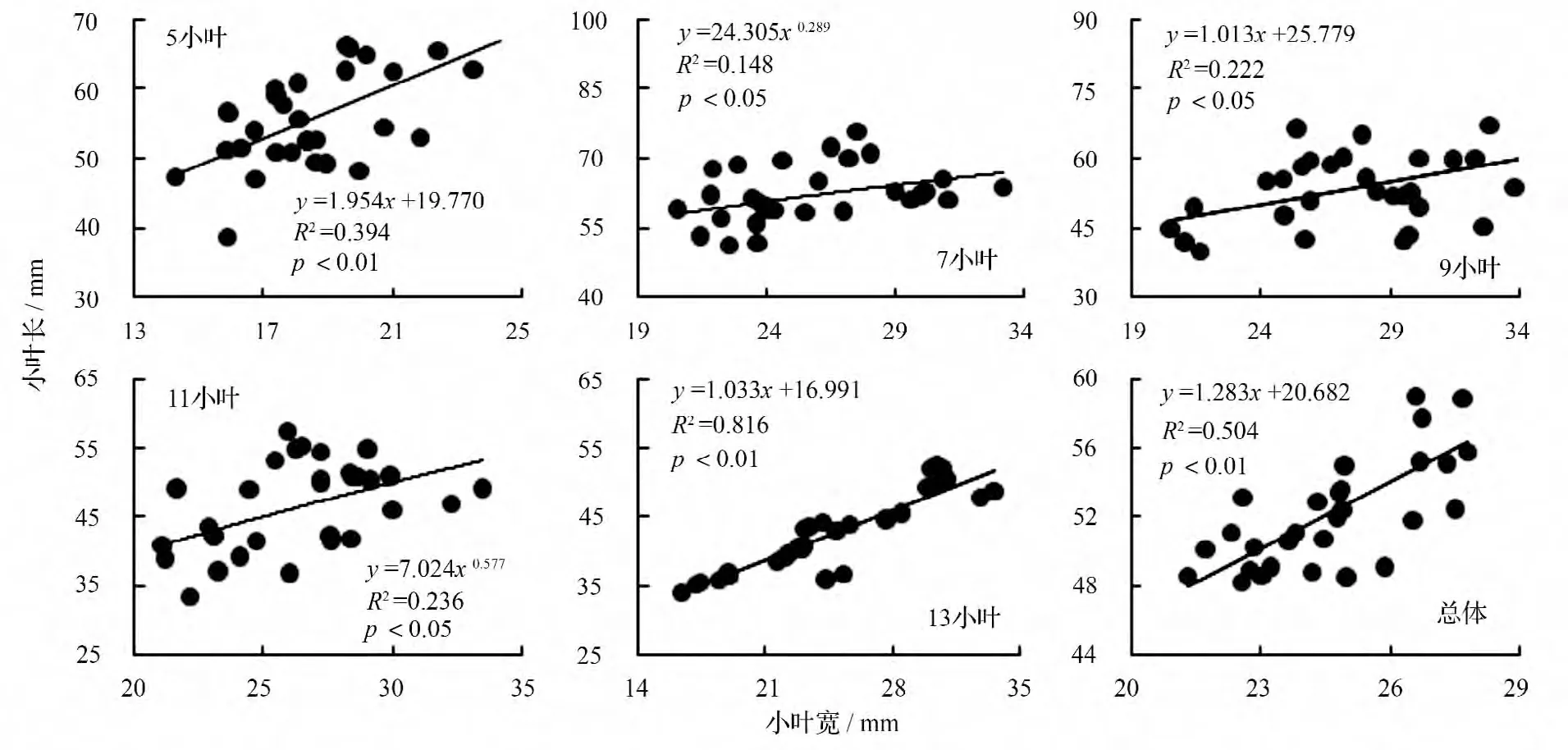
图3 小叶长与小叶宽的观测值及拟合曲线
植物个体异速生长或同速生长可看做是由物种的遗传性所决定的一个稳定特性[25],反映了生物体不同方面性状的相关程度,是生物体不同构件对环境选择压力采取的生态对策方式[26]。不同小叶白蜡复叶各构件间的生长绝大部分是异速生长型,和植物不同构件之间存在着异速生长现象是比较普遍的规律[27]相符。表明小叶白蜡复叶的生长,在具有较大可塑性的同时又具有一定的遗传稳定性,在长期的环境胁迫下已经形成了独特的生长模式。小叶白蜡复叶各构件的异速生长基本上可以用对数函数或指数函数作为定量描述模型,较好地反映植物往往朝着最佳的异速生长曲线进化策略[11]。
如果把异速生长或同速生长规律看作是由遗传因子控制的,用拟合方程的判定系数R2来估计,而随机环境影响用1 -R2来估计[13,28],则小叶白蜡5 种复叶在复叶长与叶轴长的异速生长中,有55.6%~90.6%是由遗传因子控制的,有9.4%~44.4%是受随机环境影响的;复叶长与复叶宽有53.3%~66.5%是由遗传因子控制的,有33.5%~46.7%是受随机环境影响的;小叶长与小叶宽有14.8%~81.6%是由遗传因子控制的,有18.4%~85.2%是受随机环境影响的。相比之下,在复叶长与叶轴长的相对生长中,受遗传因子的控制力较大;在复叶长与复叶宽、小叶长与小叶宽的相对生长中,遗传因子和随机环境因子均可能成为控制因子。
小叶白蜡复叶构件长或宽的标准差均远小于其平均值,表明复叶各构件在样本内比较整齐。大多数复叶构件差异显著,揭示不同小叶白蜡复叶具有不同的生长策略。复叶构件的生长既有异速生长,又有同速生长,且拟合方程的相关系数R2值有较大的差异,又揭示小叶白蜡个体间具有长期的遗传差异,多世代相传后,其个体或构件表现出多种体态。可据小叶白蜡不同类型复叶构件的性状差异,分别选作为育种目标,以更好地利用这一野生植物资源。
[1] 中国科学院中国植物志编辑委员会.中国植物志:61 卷[M].北京:科学出版社,1992:39.
[2] Bradshaw A D.Evolutionary significance of phenotypic plasticity in plant[J].Advances in Genetics,1965,13:115 -155.
[3] Schlichting C D.The evolution of phenotypic plasticity in plants[J].Annual Review of Ecology and Systematics,1986,17:667 -693.
[4] Sultan S E.Phenotypic plasticity in plants:a case study in ecological development[J].Evolution & Development,2003,5:25 -33.
[5] Pigliucci M,Kolodyska A.Phenotypic plasticity and integration in response to flooded conditions in natural accessions of Arabidopsis thaliana (L.)Heynh (Brassicaceae)[J].Annals of Botany,2002,90(2):199 -207.
[6] Sultan S E.Phenotypic plasticity and plant adaptation[J].Acta Botanica Neerlandica,1995,44:363 -383.
[7] Abulfaith H A,Bazzaz F A.The biology of Ambrosia trifida L.IV.demography of plants and leaves[J].New Phytol,1980,84:107-111.
[8] Pigliucci M,Schlichting C D.Ontogenic reaction norms in Lobelia-siphilitica (Lobeliaceae):Response to shading[J].Ecology,1995,76:2134 -2144.
[9] Bell D L,Sultan S E.Dynamic phenotypic plasticity for root growth in Polygonum:A comparative study[J].American Journal of Botany,1999,86(6):807 -819.
[10] 李斌,顾万春,卢宝明.白皮松天然群体种实性状表型多样性研究[J].生物多样性,2002,10(2):181 -188.
[11] 陆霞梅,周长芳,安树青,等.植物的表型可塑性、异速生长及其入侵能力[J].生态学杂志,2007,26(9):1438 -1444.
[12] 杨允菲,张宝田.松嫩平原草甸生境甘草种群生殖构件表型可塑性及变化规律[J].草业学报,2006,15(2):14 -20.
[13] 张维,赵玉,张相锋,等.新疆伊犁野核桃复叶的表型变异及生长规律[J].东北师大学报:自然科学版,2011,43(1):113-117.
[14] 佐艳,古丽孜亚.小叶白蜡生物学特性及栽培技术[J].农村科技,2008(7):78.
[15] 李素琼,史开奇,钟宁.小叶白腊综合性状测定[J].新疆农业科学,2001,38(3):145 -146.
[16] 谭向峰,郭霄,杜宁,等.黄栌和小叶白蜡的光合特征研究[J].山东林业科技,2010(6):1 -4,14.
[17] 蒋永喜,王少山,贺志强.小叶白蜡畸果病[J].新疆农业科学,1994(4):159 -160.
[18] 王炳举,王冬良,杨玲.小叶白蜡种子发芽试验研究[J].石河子大学学报:自然科学版,2002,6(1):31 -33.
[19] 阎洁,黄文华,潘鼎元.新疆小叶白蜡的翅果成分分析(简报)[J].石河子大学学报:自然科学版,1996(1):12.
[20] 郑健,郑勇奇,刘玉艳.野生花卉小叶白蜡引种栽培研究[J].河北农业大学学报,2007,30(6):46 -49.
[21] Pigliucci M.Phenotypic plasticity[M].Baltimore:John Hopkins University Press,2001.
[22] 张维,任艳利,赵玉,等.新疆野核桃不同小叶数复叶构件生物量可塑性及分配规律[J].东北林业大学学报,2012,40(7):37 -40.
[23] 张维,罗新泽,张娜,等.新疆野核桃种子的表型变异及生长特征[J].生态学杂志,2013,32(9):2281 -2288.
[24] Bacilieri R,Ducousso A,Kremer A.Genetic,morphological,ecological and phenological differentiation between Quercus petraea (Matt.)Liebl.and Quercus robur L.in a mixed stand of northwest of France[J].Silvae Genetica,1995,44:1 -10.
[25] Menges E S.Biomass allocation and geometry of the clonal forest herb,Laportea canadensis:Adaptive response to the environment or allometric constraints[J].American Journal of Botany,1987,74:551 -563.
[26] Primack R B.Relationships among flowers,fruits and seeds[J].Annual Review of Ecology and Systematics,1987,18:409 -430.
[27] Sun S C,Jin D M,Shi P L.The leaf size-twig size spectrum of temperate woody species along an altitudinal gradient:An invariant allometric scaling relationship[J].Annals of Botany,2006,97:97 -107.
[28] 杨允菲,李建东.松嫩平原不同生境芦苇种群分株的生物量分配与生长分析[J].应用生态学报,2003,14(1):30 -34.
猜你喜欢
昆明医科大学学报(2022年1期)2022-02-28
园艺与种苗(2022年1期)2022-02-23
昆明医科大学学报(2021年8期)2021-08-13
昆明医科大学学报(2021年6期)2021-07-31
中国计量大学学报(2019年2期)2019-08-13
山东林业科技(2019年2期)2019-06-03
山东林业科技(2019年2期)2019-06-03
花卉(2016年12期)2016-12-10
东北林业大学学报(2016年6期)2016-07-15
西南医科大学学报(2015年1期)2015-08-22
