
火棘幼苗生理生化特性对干旱胁迫的响应1)
2014-03-08 05:44肖姣娣
东北林业大学学报 2014年11期
肖姣娣
(娄底职业技术学院,娄底,417000)
责任编辑:任 俐。
随着全球气候变暖的日益加剧,干旱胁迫已经成为限制植物赖以生存、生长和分布的重要环境因子之一。而植物为了适应这种逆境,体内会发生一系列的生理生化反应,从而调动其防御与适应机制。但不同植物对干旱的抵抗能力与适应能力是不同的,由此而引起的相关反应机制与特性亦不同。因此,有针对性地开展植物对干旱胁迫的适应机理及抗旱机制研究已成为植物生理生态学研究的重点课题之一。研究结果可以为进一步明确各种植物的抗旱生理生化机理提供一定理论依据,对于筛选出各地优良节水抗旱植物也具有重要的意义。
火棘(Pyracantha fortuneana),为蔷薇科火棘属常绿灌木或小乔木,高可达4 m;主要分布于我国黄河以南及广大西南地区;可采用播种、扦插和压条法繁殖;其树形优美,枝密生有短棘,夏有繁花,秋有红果(红果药食兼用,故火棘又名吉祥果,救兵粮)且果实存留枝头甚久,是一种少见的花叶果皆具较高观赏价值的常绿植物;因其适应性强,耐修剪,喜萌发,耐贫瘠,可广泛应用于城乡园林绿化与美化,既可点缀于庭园深处、风景林地、草地边缘、园路转弯处体现自然野趣,也可布置于城市道路绿带滤尘吸毒;对矿区植被恢复与山区石漠化治理也不愧为一种良好的植物。到目前为止,对火棘的研究主要集中在育苗技术、栽培管理、开发利用等方面[1-3],不同强度干旱胁迫对火棘幼苗的影响尚未见报道。本研究通过采用盆栽人工控制水分的方法,对0.5a 火棘实生苗开展不同强度干旱胁迫处理的生理生化影响模拟试验,旨在分析探讨火棘幼苗对土壤水分干旱胁迫的适应机制与生理生化响应,以期为火棘的科学灌溉与合理利用及大面积绿化推广提供理论指导与技术支撑。
1 材料与方法
供试材料为0.5 a 火棘实生幼苗。2010年12月上旬在娄底市园林处乡土苗圃基地火棘苗木生产区选择生长健壮、无病虫害的枝条进行采种,并用塑料袋装好带回娄底职业技术学院植物与植物生理实验室堆沤3 d 闻到有腐烂味后用手搓擦果实成泥浆状,用清水中反复搅动漂洗数次后去除悬浮于上部的果皮、果肉和不饱满的秕种,留下饱满度与纯净度较高的火棘种子摊开充分阴干后,按V(种子)∶ V(细沙)=1∶ 3 混合(细沙湿度为手握成团,松手即散,稍加多菌灵粉剂)层积于瓦盆内备用。2011年1月25日在娄底职业技术学院农林实训基地搭简易小拱棚进行播种育苗,15 d 左右即相继出苗了,继续培育至3月1日揭膜露地炼苗,3月25日选择长势较好且苗高在8 cm 左右的100 株上盆到素烧泥瓦盆中进行露地盆栽培育(盆上口内径28.5 cm,下口内径24.0 cm,盆高15.0 cm,盆底内径17.0 cm),每盆1 株。盆土为混合基质(V(腐殖质土)∶ V(有机肥)∶ V(河沙)=2∶ 1∶ 1,适当加3%过磷酸钙)。土壤基本理化性质为pH 值6.6,土壤有机质、水解氮、速效磷和速效钾质量分数分别为113.4 g·kg-1,85.6、24.2、54.5 mg·kg-1,土壤相对含水量70%,用电子天平准确称量保证每盆基质质量一致。2011年5月25日(即盆栽正常培养2 个月后)从盆栽的100 盆火棘苗中按统一规格标准(平均苗高(30.00 ±1.00)cm,平均地径(0.45 ±0.50)cm,平均冠幅(6.50±1.00)cm)选择60 盆(每盆1 株)进行干旱胁迫试验。
通过单因素随机区组设计,将按统一规格标准选出的60 盆火棘苗随机分为4 个处理组(即每15盆为1 个处理组),4 个处理组分别代表4种不同强度的水分供应梯度,土壤相对含水量依次为75%(CK,对照组)、55%(LS,轻度干旱组)、35%(MS,中度干旱组)、15%(HS,重度干旱组)。土壤相对含水量,即土壤含水量占田间持水量的百分率。试验过程中通过自然失水达到预定水分胁迫条件后,采用称质量法人工控制土壤相对含水量,每天19:00对所有试验盆栽苗称质量1 次,用量杯按计算的补水量补充当天消耗的水分,使土壤含水量始终控制在设定范围内,胁迫处理持续60d 带回实验室测定各项生理生化指标。
叶绿素质量分数通过80%的丙酮提取后采用分光光度计法测定;可溶性糖质量分数采用蒽酮法测定;游离脯氨酸质量分数通过磺基水杨酸提取后采用酸性茚三酮显色法测定;相对电导率采用电导率仪法测定;丙二醛(MDA)质量摩尔浓度采用硫代巴比妥酸法(TBA)测定;过氧化物酶(POD)活性采用愈创木酚显色法测定;超氧化物歧化酶(SOD)活性采用氮蓝四唑(NBT)还原法测定。以上各项指标具体测定方法主要参照张高俊凤[4]的方法。每个指标均重复测定3 次,于测定当天09:00—10:00,选取各处理组相同部位的功能叶数片进行。
本试验属于单因素(水分)多水平试验,对取得的所有试验数据采用DPS V6.55 统计分析软件进行数据统计和方差分析,各处理平均数之间的多重比较采用Duncan 法(即SSR 新复极差测验法)进行显著性检验,用Excel 2003 软件制作数据趋势图。
2 结果与分析
2.1 干旱胁迫对火棘幼苗叶片叶绿素质量分数的影响
从表1可以看出,随着干旱胁迫强度的加大,火棘幼苗叶绿素质量分数总体呈现出下降趋势。轻度干旱胁迫(LS)与对照(CK)相比差异不显著,而中度干旱(MS)和重度干旱(HS)均显著低于对照(CK),特别是在HS 水分管理条件下,火棘幼苗的叶绿素质量分数急剧减少,只有CK 的59.39%,说明重度干旱(HS)已使火棘幼苗叶绿素合成严重受阻。
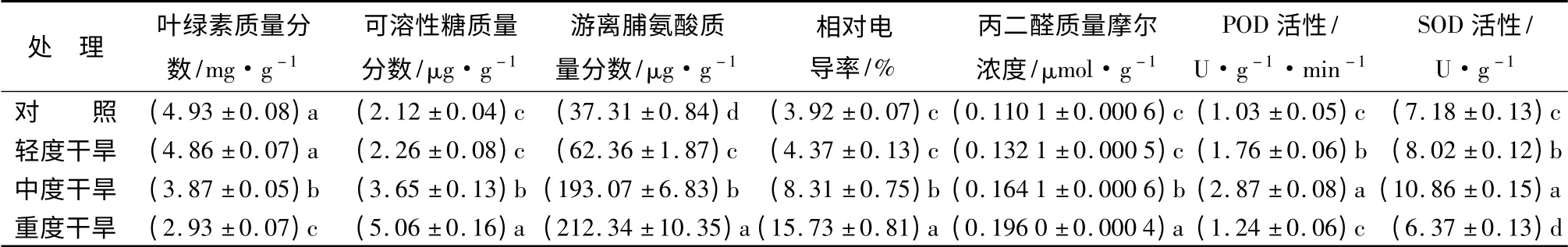
表1 干旱胁迫下火棘幼苗生理生化指标的变化
2.2 干旱胁迫对火棘幼苗叶片可溶性糖与游离脯氨酸质量分数的影响
可溶性糖质量分数:干旱胁迫条件下火棘叶片中可溶性糖质量分数的变化如表1所示。随着干旱胁迫的加重,可溶性糖质量分数呈现增加趋势。其中轻度干旱(LS)增加缓慢,只比对照(CK)高出6.19%,与CK 差异不显著,而中度干旱(MS)和重度干旱(HS)则分别比对照(CK)高出64.70%、138.68%,与CK 差异显著。说明在水分严重缺乏时,火棘可通过增加可溶性糖质量分数缓解、抵抗干旱对其造成的伤害。
游离脯氨酸质量分数:如表1所示,随着干旱胁迫的加重,火棘叶片中游离脯氨酸质量分数增加非常明显。重度干旱(HS)条件下达到最高值212.34 μg·g-1,轻度干旱(LS)、中度干旱(MS)和重度干旱(HS)时脯氨酸质量分数分别是对照(CK)的1.67 倍、5.17 倍、5.69 倍;且在CK、LS、MS、HS 条件下火棘幼苗叶片中游离脯氨酸质量分数两两之间差异均达到显著水平。说明产生大量的游离脯氨酸是火棘幼苗应对干旱的一种保护响应策略。
2.3 干旱胁迫对火棘幼苗叶片相对电导率和丙二醛质量摩尔浓度的影响
相对电导率:由表1可知,随着干旱胁迫强度的加深,火棘叶片相对电导率不断上升。其中,中度干旱(MS)和重度干旱(HS)胁迫下火棘叶片的电导率均显著高于对照(CK),分别是对照(CK)的2.12倍、4.01 倍,而轻度干旱(LS)的电导率稍高于对照(CK),但差异不显著。
丙二醛质量摩尔浓度:随着干旱胁迫强度的加深,火棘幼苗叶片丙二醛(MDA)质量摩尔浓度不断增加(表1),同时增速比较均匀。轻度干旱(LS)时,MDA 质量摩尔浓度与对照(CK)差异不明显,但在中度干旱(MS)和重度干旱(HS)条件下分别比对照(CK)高出49.05%、78.01%,差异达到显著水平。
2.4 火棘幼苗叶片过氧化物酶和超氧化物歧化酶活性对干旱胁迫的响应
过氧化物酶活性:从表1可看出,火棘幼苗叶片过氧化物酶(POD)活性对干旱胁迫的响应表现出了先增加后下降的变化趋势,同时增加的速度与下降的速度均比较快。轻度干旱(LS)、中度干旱(MS)时叶片的POD 活性分别是对照(CK)的1.71倍、2.78 倍,与CK 差异明显;而重度干旱(HS)与轻度干旱(LS)、中度干旱(MS)相比,POD 活性显著减弱而与他们差异显著,但重度干旱(HS)时其活性仍然稍高于对照(CK),但两者差异不显著,这说明即使在严重干旱胁迫条件下火棘还能维持较高的POD 活性,抗旱耐旱性较强。
超氧化物歧化酶活性:随着干旱胁迫的加强,火棘幼苗叶片超氧化物歧化酶(SOD)活性也呈现出先增强后减弱的变化特点(表1)。其中在中度干旱(MS)处理条件下SOD 达到最大值,说明一定程度的干旱胁迫(试验中LS、MS 处理条件下)可以使火棘幼苗叶片的SOD 活性明显增强,以应对干旱对其造成的伤害;但当干旱达到一定程度后(试验中HS处理条件下),火棘幼苗叶片SOD 活性又显著受到抑制而减弱,其活力已显著低于对照(CK),而轻度干旱(LS)与中度干旱(MS)胁迫下SOD 活性显著高于对照(CK)。
3 结论与讨论
叶绿素是植物将捕获的光能转化为化学能的重要活性物质,植物叶片中叶绿素质量分数的高低将直接影响植物光合作用的强弱与效率,进而影响植物的生长发育。本研究表明,轻度干旱胁迫下(即土壤相对含水量55%),火棘幼苗叶片中叶绿素质量分数下降不明显,而随干旱胁迫强度的加重,火棘幼苗叶片中叶绿素质量分数呈现出明显下降趋势。原因可能是因为较强的干旱胁迫条件制约了叶绿素的生物合成,加速了叶绿素酶活性的分解,迫使叶绿素质量分数下降[5]。但黄小辉等[6]研究水分胁迫对桑树幼苗生理特性的影响发现,干旱锻炼的桑树幼苗叶片的叶绿素质量分数反而提高,并认为这是植物在缺水状态下气孔关闭,胞间CO2含量降低,通过提高叶绿素质量分数来适应与抵抗干旱胁迫造成的影响。
可溶性糖与脯氨酸是植物体内重要的渗透调节物质,可缓解、抵抗不良环境对植物造成的伤害[7]。本研究表明,随干旱胁迫强度的加重,火棘叶片中可溶性糖与游离脯氨酸质量分数不断增加,特别是在重度干旱(土壤相对含水量15%)时可溶性糖与游离脯氨酸质量分数分别是对照(土壤相对含水量75%)的2.39 倍、5.69 倍。这可能是当火棘遇到严重干旱缺水的不良环境条件时,能够通过代谢活动诱导植物体内积累较多的可溶性糖与脯氨酸来增加细胞溶质的浓度,维持细胞较低的渗透势,有利于细胞或组织的吸收能力与保水能力,从而抵抗干旱胁迫带来的伤害,提高抗旱适应能力[8]。这与吴庆贵等[9]研究珙桐、邱真静等[10]研究沙拐枣对干旱胁迫的响应结果一致。
相对电导率的高低可以反映细胞膜透性的大小与膜系统稳定性的重要指标,植物在受到逆境胁迫时,首先表现为细胞膜受损,导致细胞膜透性增大,其值越大,表示细胞质膜受伤害的程度越重[11];而丙二醛(MDA)是细胞膜发生膜脂过氧化反应的产物,其质量摩尔浓度的高低常用来衡量植物在逆境胁迫中受活性氧伤害的程度[12-13]。本研究中,相对电导率与MDA 质量摩尔浓度的变化规律相类似,均表现为轻度干旱(LS)胁迫下与对照差异不明显,而中度干旱(MS)和重度干旱(HS)均显著高于对照(CK),说明细胞膜透性的变化与膜脂过氧化反应是缓慢渐进的,只有当干旱胁迫到一定程度后,细胞膜结构与功能才会受到严重的伤害,引起膜透性增大与膜脂过氧化程度的加重,从本研究结果来看,质膜透性与质膜过氧化程度之间可能存在一定相关性,与孟健男等[14]对冬小麦、史宝胜等[15]对金叶榆的研究结果相一致。
POD(过氧化物酶)和SOD(超氧化物歧化酶)是植物体内重要的抗氧化酶保护系统,其主要作用是清除植物体内产生的活性氧和自由基,避免或减轻它们对植物造成的氧化伤害[16]。本研究表明,随着干旱胁迫强度的加重,火棘幼苗叶片的POD、SOD活性均呈现出先增强后减弱的变化特点,说明当植株受到轻度干旱(LS)、中度干旱(MS)胁迫时,其体内会增强这两种保护酶活性来维持植株正常的生命活动;但POD、SOD 活性增强也是有限度的,当干旱胁迫超出了一定范围,保护酶活性就会显著受到抑制而减弱。
综上所述,火棘幼苗能够通过调节自身叶绿素合成、积累可溶性糖与脯氨酸、改变膜透性与MDA 质量摩尔浓度以及变化POD、SOD 活性等生理生化特征来适应与应对不同干旱胁迫对其生命活动与生长造成的影响,表现出一定的抗旱耐旱潜力。但植物的抗旱耐旱性是多种途径共同作用的结果,衡量植物抗旱耐旱性的指标也很多,本研究只从火棘幼苗叶片一些生理生化指标特性分析了其对干旱胁迫的响应,而其他指标特性方面还有待进一步研究与探讨。
[1] 郭红梅.火棘温棚育苗技术初探[J].青海农林科技,2013(1):59 -60,47.
[2] 曾明颖,罗学刚.火棘的开发利用[J].绵阳经济技术高等专科学校学报,1999,16(4):34 -37.
[3] 张玉芹.火棘容器育苗技术研究[J].北方园艺,2011(2):95-97.
[4] 高俊凤.植物生理学实验指导[M].北京:高等教育出版社,2006.
[5] 贾瑞丰,徐大平,杨曾奖,等.干旱胁迫对降香黄檀幼苗光合生理特性的影响[M].西北植物学报,2013,33(6):1197 -1202.
[6] 黄小辉,刘芸,李佳杏,等.水分胁迫对三峡库区消落带桑树幼苗生理特性的影响[J].林业科学,2012,48(12):122 -126.
[7] 朱广龙,韩蕾,陈婧,等.酸枣生理生化特性对干旱胁迫的响应[J].中国野生植物资源,2013,32(1):33 -37.
[8] 魏良明,贾了然,胡学安,等.玉米抗旱性生理生化研究进展[J].干旱地区农业研究,1997,15(4):68 -73.
[9] 吴庆贵,杨敬天,邹利娟,等.珙桐幼苗生理生态特性对土壤干旱胁迫的响应[J].江苏农业科学,2014,42(2):119 -122.
[10] 邱真静,李毅,种培芳.PEG 胁迫对不同地理种源沙拐枣生理特性的影响[J].草业学报,2011(3):108 -114.
[11] 兆慧茹,王丽娟,郑蕊,等.宁夏5 种抗旱性牧草与脯氨酸含量的相关性研究[J].宁夏农学院学报,2001,22(4):12 -14.
[12] 何淼,高耀辉,徐鹏飞,等.中国芒幼苗对土壤干旱胁迫的生理响应[J].安徽农业科学,2013,41(30):12125-12128,12131.
[13] 丰庆荣,徐庆华,黄有龙.土壤含水量对长白落叶松幼苗物质含量的影响[J].林业实用技术,2011(4):9 -11.
[14] 孟健男,于晶,苍晶,等.PEG 胁迫对两种冬小麦苗期抗旱生理特性的影响[J].东北林业大学学报,2011,42(1):40 -44.
[15] 史宝胜,刘冬云,张小磊,等.水分胁迫对金叶榆含水量、细胞质膜相对透性和抗氧化系统的影响[J].西北农业学报,2010,19(9):88 -92.
[16] 彭立新,李德全,束怀瑞,等.园艺植物水分胁迫生理及耐旱机制研究进展[J].西北植物学报,2002,22(5):1275 -1281.
猜你喜欢
中老年保健(2022年3期)2022-11-21
知识窗(2021年1期)2021-02-23
农村实用技术(2020年4期)2020-12-15
世界科学(2020年1期)2020-02-11
西江月(2019年1期)2019-11-20
科学与财富(2019年15期)2019-10-21
中国生物医学工程学报(2019年5期)2019-07-16
花卉(2017年6期)2017-07-31
Coco薇(2017年5期)2017-06-05
湖南林业科技(2017年1期)2017-02-06
