
辽东山区次生林土壤微生物生物量对微生物呼吸影响
2014-03-06 03:19范阿南
东北林业大学学报 2014年3期
范阿南 刘 峰
(大连市林业科学研究院,大连,116039) (内蒙古赤峰市红山区国营城郊林场)
土壤微生物生物量是指土壤中除活的植物体外体积小于5 ×103μm3的生物总量,主要包括细菌、真菌、藻类和原生动物等[1-2]。微生物生物量虽然只占土壤有机质库的很小部分,但却是控制生态系统中碳、氮和其它养分流的关键[3],影响着所有进入土壤的有机质的转化,是整个生态系统养分和能源循环的“关键”和“动力”[4]。土壤微生物生物量同时也是土壤养分转化过程中的一个重要的源和库[5]。土壤微生物生物量对环境变化敏感,能够较早地指示生态系统功能的变化[6],微生物量库的任何变化将会影响养分的循环和有效性[7]。
辽东山区的植物区系主要以长白植物区系为主,近100年来由于特殊的历史和社会原因使辽东山区的森林遭到了几次大规模的破坏,在剧烈的人为干扰下,这一地区稳定的顶级群落阔叶红松林大部分消失,形成了目前主要以柞属(Quercus)、桦属(Betura)、杨属(Populus)、槭属(Acer)等树种为主的次生林。本研究以辽东山区天然次生林为研究对象,较系统地研究了主要天然次生林植被类型在森林演替过程中的土壤微生物生物量以及土壤微生物呼吸强度,为深入了解干扰条件下土壤微生物生物量碳、氮在森林生态系统物质循环和能量流动的功能提供基础数据,为该区域的森林经营实践提供理论参考。
1 研究区概况
研究地位于中国科学院沈阳应用生态研究所清原森林生态实验站(海拔456 ~1 116 m)。气候属受季风影响的温带大陆性气候,主要土壤种类为棕色和暗棕色森林土,植物区系主要以长白植物区系为主。这一地区曾遭遇几次大规模的破坏,原有顶级群落阔叶红松林大部分消失,现有植被以次生林为主。胡理乐等[8]将辽东山区天然次生林划分为蒙古栎、胡桃楸、杂木林、水曲柳和花曲柳5 个主要群落类型,本研究以其中的蒙古栎、胡桃楸和杂木林3个类型为研究对象,试验地林分特征见表1。
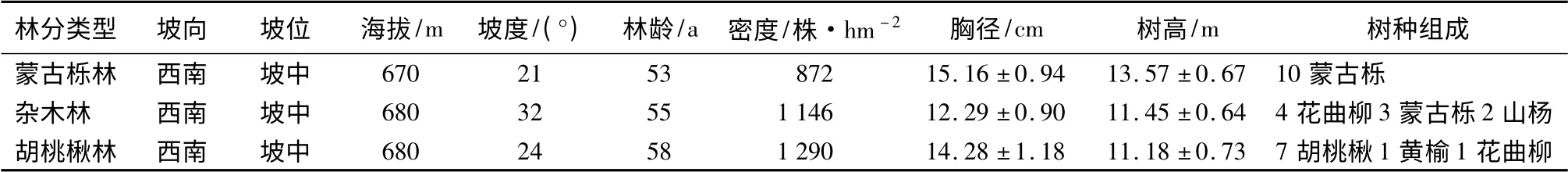
表1 辽东山区次生林样地基本特征
2 材料与方法
试验地设置:选取具有代表性的蒙古栎林、杂木林、胡桃秋林林分,每个林分内设置30 m ×30 m 固定样地,试验在20 m ×20 m 区域内进行,10 m 作为缓冲区。对林地内胸径大于4 cm 的树木进行每木检尺,测量树高和胸径。
土壤微生物呼吸测定:土壤表面CO2通量采用LI-COR 公司生产的LI-6400 便携式测定系统连接的土壤呼吸室(LI-Cor Inc.,Lincoln,NE,USA)直接测定。土壤微生物异养呼吸(RH)采用挖壕法[5]间接测定。挖壕6 个月后,测定壕内土壤呼吸速率,测得的挖壕样方内的CO2通量即为土壤微生物呼吸速率。
土壤微生物生物量土样采集、制备:每月土壤呼吸测定后,在每个土壤呼吸观测点附近用钻土芯法采集土样。土钻直径为4 cm,每个采样点分0 ~10、>10 ~20、>20 ~30 cm 3 个层次分别收集,每个观测点选4 个采样点,将采集到的每个层次的4 个重复混合作为一个样品装入密封袋中,回实验室后迅速过筛。过筛后的土壤(D <2 mm)用作土壤微生物量测定。
微生物生物量碳、氮测定:氯仿熏蒸浸提[9],用碳、氮自动分析仪(Multi N/C3000,Germany)测定碳、氮质量分数。
数据处理与统计方法:采用Excel(2003)和SPSS(11.5)对试验数据进行统计、分析和制图。
3 结果与分析
3.1 不同林分土壤微生物生物量碳的动态变化
蒙古栎林、杂木林和胡桃楸林土壤微生物生物量碳质量分数均具有明显的季节变化规律,生长季内呈单峰趋势(表2)。4月份3 个群落土壤微生物生物量碳最低,5、6月份逐渐升高,至7月份升至整个生长季的最高值,随后再逐渐下降。整个生长季,蒙古栎林微生物生物量碳大于杂木林,但只在10月份差异显著,胡桃楸林的微生物生物量碳最小,但也只在4、5、7、9月份与其它2 种群落差异显著。

表2 辽东山区不同林分土壤微生物生物量碳、氮季节变化 mg·kg -1
3.2 不同林分土壤微生物生物量氮的动态变化
蒙古栎林、杂木林、胡桃楸林土壤微生物生物量氮质量分数均表现明显的季节变化,均呈单峰趋势(表2)。各群落土壤微生物生物量氮质量分数在4月份最低,5、6月份逐渐增加,蒙古栎林和杂木林在7月份微生物生物量氮最高,而胡桃楸林在8月份最高,随后各群落土壤微生物生物量氮质量分数均逐渐降低。生长季土壤微生物生物量氮质量分数大小顺序依次为蒙古栎林、杂木林、胡桃楸林。4月份3 个群落微生物生物量氮质量分数分别为5.87、4.24、3.78 mg/kg,蒙古栎林比杂木林高38%,差异不显著,胡桃楸林最低,只是蒙古栎和杂木林的64%和89%。5、6月份3 个群落微生物生物量氮质量分数间差异不显著。7月份3 个群落微生物生物量氮质量分数分别为241.62、184.89、76.71 mg/kg,蒙古栎林比杂木林高31%,但差异不显著,胡桃楸林最低,是蒙古栎林和杂木林的32%和41%。8月份3 个群落微生物生物量氮质量分数依次为215.72、146.72、123.37 mg/kg,蒙古栎林分别是杂木林和胡桃楸林的1.47、1.75 倍,杂木林比胡桃楸林高,但二者差异不明显。9月份,3 个群落微生物生物量氮质量分数分别为180.20、144.74、71.86 mg/kg,蒙古栎林比杂木林高,但二者差异不明显,胡桃楸林微生物生物量氮质量分数最低,明显低于蒙古栎林和杂木林,仅占二者的40%和50%。10月份3 个群落的微生物生物量氮质量分数分别为100.41、36.25、21.78 mg/kg,相对于9月份降低较明显,蒙古栎林生物量是杂木林和胡桃楸林的2.77、4.61 倍,杂木林微生物生物量氮质量分数虽然高于胡桃楸林,但二者差异不明显。
3.3 不同林分土壤微生物呼吸速率的变化
生长季蒙古栎林、杂木林、胡桃楸林土壤微生物呼吸速率及变化规律不同(图1)。土壤微生物呼吸速率由大到小为蒙古栎林、杂木林、胡桃秋林;蒙古栎林和杂木林土壤微生物呼吸速率最低值出现在4月份,分别为0.74、0.66 g·m-2·d-1,而胡桃秋林土壤微生物呼吸速率则在生长季结束的10月份达到最低,为0.84 g·m-2·d-1;8月蒙古栎林和胡桃秋林土壤微生物呼吸速率达到峰值,分别为2.77、1.83 g·m-2·d-1,而杂木林群落土壤微生物呼吸速率则在6月份就已达到最大,为1.98 g·m-2·d-1。
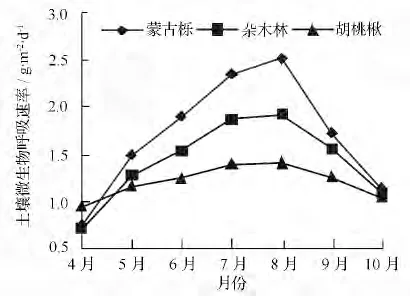
图1 辽东山区不同林分生长季土壤微生物呼吸速率的变化
3.4 土壤微生物生物量碳、氮与土壤微生物呼吸的关系拟合
对土壤微生物呼吸与土壤微生物生物量碳、氮进行回归分析,结果见表3。回归分析表明,蒙古栎林、杂木林和胡桃楸林土壤微生物呼吸与土壤微生物生物量碳、氮均具有线性关系。土壤微生物是所有进入土壤的有机质的分解者和转化者,土壤微生物碳是活的有机质部分,它的变化与土壤生物活性密切相关,直接或间接影响土壤呼吸的强弱。Scott-Denton 等[10]认为,土壤呼吸速率与微生物生物量碳关系密切;Liu 等[11]对长白山典型森林生态系统土壤呼吸进行研究表明,土壤呼吸和土壤微生物生物量碳之间呈正相关关系,但未达到显著水平;而Sato 等[12]的研究却发现二者之间没有关系。目前有关土壤呼吸与微生物生物量氮之间的研究并不多。辽东山区次生林蒙古栎林、杂木林和胡桃楸林土壤微生物呼吸与土壤微生物生物量氮的相关性好于与土壤微生物生物量碳的相关性。蒙古栎林和胡桃楸林土壤微生物呼吸与微生物生物量碳之间的决定系数分别为0.633 4 和0.669 0,关系显著(P <0.05),杂木林微生物呼吸与微生物生物量碳之间具有极显著的线性关系(R2=0.852 3,P <0.01);蒙古栎林、杂木林和胡桃楸林土壤微生物呼吸与微生物生物量氮之间线性回归决定系数分别为0.824 4、0.907 9 和0.871 8,均达极显著水平(P <0.01)。说明土壤微生物生物量大小决定了土壤微生物呼吸的强弱。
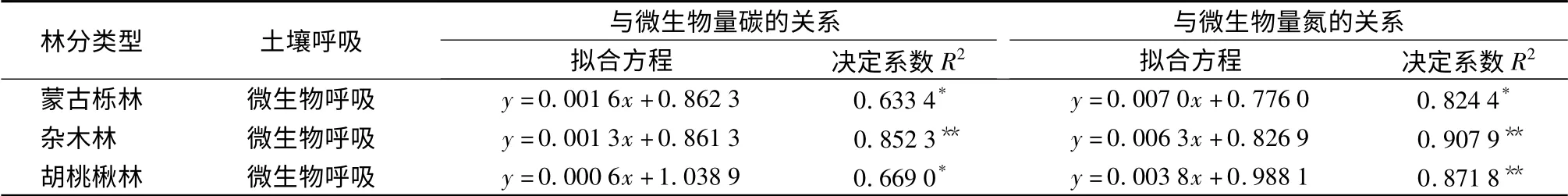
表3 辽东山区林分土壤微生物呼吸与土壤微生物生物量碳、氮关系的拟合方程
4 结论
次生林群落土壤微生物生物量碳、氮均具明显的季节变化,生长季内呈单峰曲线变化,基本于7、8月达到最大值;生长季内,蒙古栎林微生物生物量碳质量分数最高,胡桃楸林最低,彼此间差异不显著。土壤微生物生物量氮质量分数大小顺序与微生物生物量碳一致,胡桃楸林土壤微生物生物量氮质量分数明显低于蒙古栎林和杂木林。蒙古栎林和胡桃楸林土壤微生物呼吸与微生物生物量碳间决定系数分别为0.633 4 和0.669 0,关系显著(P <0.05),杂木林微生物呼吸与微生物生物量碳之间具有极显著的线性关系(R2=0.852 3,P <0.01);蒙古栎林、杂木林和胡桃楸林土壤微生物呼吸与微生物生物量氮之间线性关系极显著,决定系数分别为0.824 4、0.907 9 和0.871 8(P <0.01)。
[1] 刘海琴,王志明,朱培立,等.施加有机肥对土壤微生物量及其周转的影响[J].江苏农业科学,2002(5):69-71.
[2] 王岩,沈其荣,史瑞和,等.土壤微生物量及其生态效应[J].南京农业大学学报,1996,19(4):45-51.
[3] Jenkinson D S.The determination of microbial biomass carbon and nitrogen on soils[M]//Wilson J R.Advances on nitrogen cycles in agricultural ecosystems.Wallingford:C A B International,1988:368-386.
[4] 李延茂,胡江春,汪思龙,等.森林生态系统中土壤微生物的作用与应用[J].应用生态学报,2004,15(10):1943-1946.
[5] 周建斌,陈竹君,李生秀.土壤微生物量氮含量、矿化特性及其供氮作用[J].生态学报,2001,21(10):1718-1725.
[6] Anderson T H,Domsch K H.The metabolic quotient for CO2(qCO2)as a specific activity parameter to access the effects of environmental conditions such as pH,on the microbial biomass of forest soils[J].Soil Biology and Biochemistry,1993,25(3):393-395.
[7] Roy S,Singh J S.Consequences of habitat heterogeneity for availability of nutrients in a dry tropical forest[J].Journal of Ecology,1994,82(3):503-509.
[8] 胡理乐,毛志宏,朱教君,等.辽东山区天然次生林的数量分类[J].生态学报,2005,25(11):2848-2854.
[9] Wu J,Joergensen R G,Pommerening B,et al.Measurement of soil microbial biomass by fumigation-extraction-an automated procedure[J].Soil Biology and Biochemistry,1990,22(8):1167-1169.
[10] Scott-Denton L E,Sparks K L,Monson R K.Spatial and temporal controls of soil respiration rate in a high-elevation,subalpine forest[J].Soil Biology and Biochemistry,2003,35(4):525-534.
[11] Liu Ying,Han Shijie,Zhou Yumei,et al.Contribution of root respiration to total soil respiration in a Betula ermanii-Dark coniferous forest ecotone of the Changbai Mountains,China[J].Pedoshpere,2005,15(4):448-455.
[12] Sato A,Seto M.Relationship between rate of carbon dioxide evolution,microbial biomass carbon,and amount of dissolved organic carbon as affected by temperature and water content of a forest and an arable soil[J].Community Soil Science and Plant Analysis,1999,30(19/20):2593-2605.
猜你喜欢
阅读与作文(小学高年级版)(2019年8期)2019-10-16
蒙古学问题与争论(2019年0期)2019-03-29
电影(2018年11期)2018-11-24
西域历史语言研究集刊(2018年0期)2018-11-09
现代农业研究(2017年11期)2018-01-12
辽宁林业科技(2017年4期)2017-06-22
辽宁林业科技(2017年4期)2017-06-22
草原歌声(2017年1期)2017-04-23
学与玩(2017年4期)2017-02-16
现代园艺(2017年20期)2017-02-03
