
不同空气相对湿度对槭叶草生长及光合生理特性的影响1)
2014-03-06 03:19张爽董然董妍赵超
东北林业大学学报 2014年3期
张 爽 董 然 董 妍 赵 超
(吉林农业大学,长春,130118) (长春友谊公园管理所) (吉林农业大学)
槭叶草(Mukdenia rossii (Oliv.)Koidz)是虎耳草科(Saxifragaceae)槭叶草属(Mukdenia)多年生草本植物,在我国主要分布于吉林省长白山一带,属吉林省重点保护野生植物[1];其叶形独特,掌状5 ~7中裂至深裂,秋叶变红,株高20 ~30 cm,早春开花,花色洁白,极具观赏价值,是优良的园林地被植物。但目前槭叶草并未在园林绿化中大规模推广应用,这是因为野生植物需要良好的适应性才能在园林中各种立地条件下正常生长发育,发挥出最佳的美化和生态作用,槭叶草也不例外。
目前,对槭叶草的研究主要集中在药理药效及繁殖技术等方面[2-3],对其在园林中的应用研究较少,而对其生态适应性的研究更未见报道。虽然马全等[4]通过其原生境判断槭叶草在园林绿化中适于栽植于假山、溪水的绿化区、绿化带,但缺乏理论依据。宫敬利[5]在对槭叶草原生境的调查中发现,采样地的年相对湿度70%左右,并且在周围植被较好、空气湿度较大的的环境条件下槭叶草生长最好,而在炎热干燥的天气其叶片常过早老化。可见,空气相对湿度直接影响着槭叶草生长发育和观赏效果。所以,本试验采用自动控制空气相对湿度的方法,研究不同空气相对湿度对槭叶草生长及光合作用的影响,从生长和光合特性两个方面探寻其最适空气湿度范围,以期为更好地发挥其观赏价值,丰富东北地区园林绿化中早春植物种类,确定其应用范围提供理论支持和实践依据。
1 材料与方法
试验地位于长春市吉林农业大学园林试验基地,属大陆性季风气候,春季干燥多风,夏季温热多雨,秋季晴朗温差较大,冬季漫长寒冷。年平均气温为4.8℃,夏季7月份最高温度39.5 ℃,冬季1月份最低温度-39.8 ℃,年平均降水量为567.0 mm,夏季降水量占全年降水量的60%以上,是降水量最丰富时期。
供试材料为长白山野生槭叶草的3年生种子苗,2013年3月10日将具有相同芽数的根茎移栽到直径为20 cm 的塑料盆中,盆内基质V(园土)∶ V(沙子)∶ V(草炭)=2∶ 1∶ 1,置于园林试验基地的温室内缓苗,31 d 后进行不同湿度处理,期间根据温室的条件进行正常肥水管理(0.3%尿素和磷酸二氢钾,2 次·月-1)。
2013年4月10日选取株高、冠幅大致相同的槭叶草移入不同空气相对湿度处理下的塑料棚(2 m×3 m)中,塑料棚内将湿度探测头分别设定空气相对湿度(RH)为45%(CK)、65%、75%、85%(采用加湿器与湿度探测器连接的XTM9001 自动化控制系统,使得棚内湿度始终控制在设定值范围);采用随机区组设计,每个处理10 株,3 次重复;试验于6月10日结束,处理时间60 d。处理期间每天早晚各进行一次排风换气,每3 d 浇1 次水。
生长指标和叶绿素质量分数的测定:试验处理前和处理结束时分别测定1 次株高、叶长、叶宽、冠面积,并记录叶片数。冠面积=0.25π ×冠幅长×冠幅宽[6];叶面积、比叶质量参考丁爱萍等[7]的方法。叶绿素质量分数,试验结束时取生长方向相同的新生成熟叶片,参考李合生[8]的方法测定。
光合特性的测定:2013年6月14日,选择晴朗无云的天气,利用美国PPSystems 公司生产的CIRAS-2 型便携式光合仪进行测定槭叶草的光合值。选取长势、朝向一致的供试槭叶草功能叶片,每个处理选3 株,每株测定3 片叶,共9 次重复。测定时间为06:00—18:00,每隔2 h 测定1 次;测定项目为净光合速率(Pn)、胞间CO2摩尔分数(Ci)、气孔导度(Gs)、蒸腾速率(Tr)等,计算水分利用率(WU。E),WU。E=Pn/Tr和气孔限制值(Ls),Ls= 1- Ci/Ca。(其中,Ca为叶外空气的CO2摩尔分数)。按照梁宇等[9]的方法,计算净光合累积值和蒸腾累积值。
数据分析:使用DPSv7.05 软件对数据进行统计分析,采用Duncan 新复极差法进行显著性差异分析,采用SPSS17.0 进行相关性分析。
2 结果与分析
2.1 不同空气湿度对槭叶草生长的影响
由表1可以看出,不同RH对槭叶草的生长影响较大。随着RH的升高新生叶片数增多,植株高度、冠面积、叶面积的净增长率及比叶质量也明显增大。85%RH下各生长指标达到最大值,且其余各处理与45%RH下各指标间差异显著。
2.2 不同空气湿度对槭叶草叶绿素质量分数的影响
由表2可以看出,不同RH处理对槭叶草叶片中叶绿素a 的影响较大,对叶绿素b 的影响较小,从而导致叶绿素(a +b)和叶绿素a/b 的变化全趋势与叶绿素a 相同。各RH处理下的叶绿素质量分数由大到小依次为:75%RH、85%RH、65%RH、45%RH,且45%RH与其他3 个处理间差异显著。

表1 不同空气湿度对槭叶草生长的影响

表2 不同空气湿度对槭叶草光合色素质量分数的影响
2.3 不同空气湿度对槭叶草光合生理参数日变化的影响
2.3.1 环境因子日变化
由表3可知,光合有效辐射和空气温度的日变化趋势一致,从06:00 开始逐渐上升,12:00 时达到最大值,此后则不断下降。外界CO2摩尔分数则呈先下降后上升的趋势,06:00 时最高,此后逐渐下降,至16:00 时逐渐升高。
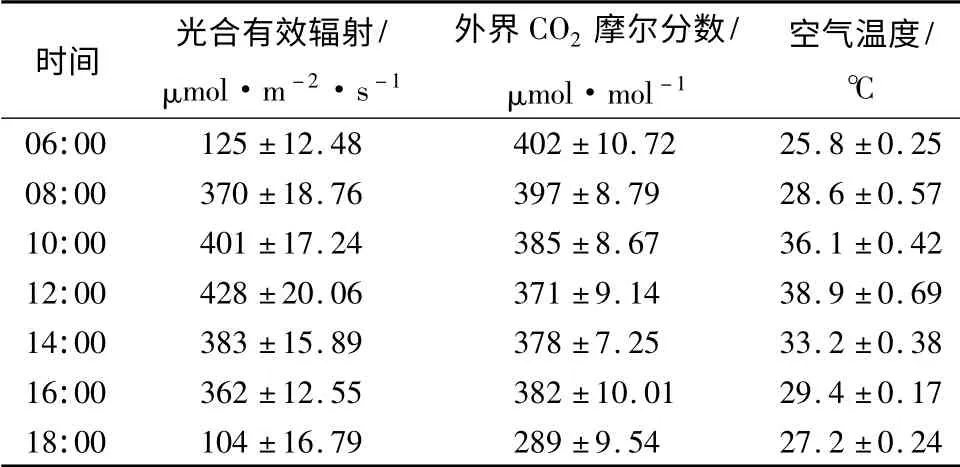
表3 环境因子日变化
2.3.2 不同空气湿度对槭叶草净光合速率日变化的影响
由表4可以看出,在45%、65%、75%RH处理下槭叶草的净光合速率(Pn)随时间变化趋势均呈“双峰”曲线;85%RH下Pn呈“单峰”曲线。不同RH处理下峰值出现的时间不同。45%RH下的两次峰值出现在08:00 和16:00,谷值出现在14:00;65%和75%RH处理下的Pn两次峰值均出现在10:00 和14:00,谷值出现在12:00,3 个处理下的槭叶草均表现出典型的光合“午休”现象,且第2 次峰值均低于第1 次峰值。而85%处理下的峰值出现在10:00,10:00 之后逐渐降低。净光合积累量随空气湿度的增加而增大,45%RH下的是65%、75%、85%RH下的53.53%、49.40%、36.08%。说明在一定范围内空气湿度的增加能够缩短槭叶草光合“午休”的时间,而较高的空气湿度能够使光合“午休”现象消失,从而有利槭叶草光合产物的积累。
2.3.3 不同空气湿度对槭叶草胞间CO2摩尔分数和气孔限制值的影响
由表4可以看出,各RH处理下槭叶草胞间CO2摩尔分数(Ci)均呈早中晚较高,其他时间段较低的变化趋势,大致呈“W”型曲线,说明槭叶草早晚和中午利用CO2的能力较低;不同RH处理下Ci由大到小依次为:85%、75%、65%、45%RH。气孔限制值的日变化趋势呈“双峰”曲线,不同空气湿度处理下峰值出现的时间不同;随空气湿度的增加Ls不断减小,且与Ci的变化趋势相反。
2.3.4 不同空气湿度对槭叶草蒸腾速率、气孔导度及水分利用率日变化的影响
由表4可以看出,各RH处理下槭叶草蒸腾速率(Tr)、气孔导度(Gs)随时间变化趋势均大致呈“双峰”曲线;45%RH下Tr的2 次峰值出现在10:00和16:00,其余各处理均出现在10:00 和14:00。各RH处理下的Gs均在06:00 最高,此后不断降低,45%RH第2 次峰值出现在16:00,其余各处理均在14:00。Tr和Gs的变化趋势与Pn相似。45%、65%、75%RH下的WU。E均在8:00 最高,85%RH在10:00 最高,此后均不断下降;45%和85%RH的第2次峰值出现在16:00,而65%和75%RH的出现在14:00。

表4 不同空气湿度对槭叶草光合参数的影响
2.4 空气湿度与各光合生理参数的关系
植物光合作用受外部环境因子和内在生理因子的双重影响。为了更好地分析空气相对湿度对槭叶草光合作用的影响,对RH与各光合生理参数进行相关性分析。由表5可以看出,RH与Pn、Gs和Tr呈显著正相关,与Ls呈极显著负相关,与Ci和WU。E无显著相关性。Pn与Ci、Gs和Tr呈显著正相关,与Ls呈显著负相关关系,与WU。E无显著相关性。
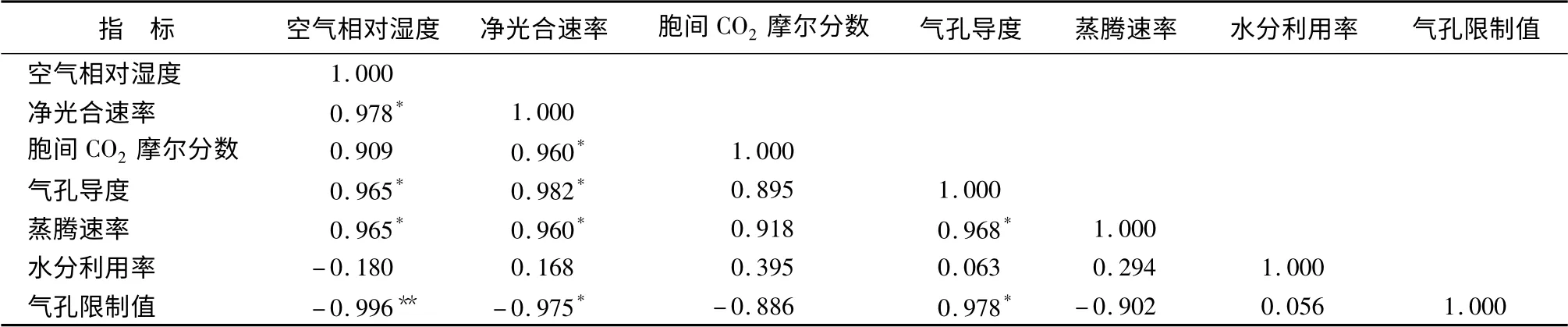
表5 空气湿度与各光合生理参数的相关性分析
3 结论与讨论
植株形态是植物适应外界环境最直观的表现形式。研究结果表明,随着空气湿度的增加,槭叶草株高、叶片数、叶面积和冠面积的净生长率均不断增大,且其余各处理与45%RH间差异显著,说明增加空气湿度能显著促进槭叶草生长。这与尤伟忠[10]对东方百合的研究结果一致。
叶绿素是参与光合作用光能吸收、传递和转化的重要色素[11]。研究结果表明,叶绿素质量分数由大到小依次为75%、85%、65%、45%RH,这与张宇等[12]的研究结果不尽相同,许大全[11]认为,空气湿度较大,植物叶绿素会产生不同程度的降解,但并不影响植物的光合作用。
在自然条件下,植物光合作用日变化曲线大致有2 种,一种是“单峰”曲线:中午净光合速率最高;第二种是“双峰”曲线,上午和下午各有一个峰值[13-14]。光合“午休”是一种比较普遍的现象,在许多植物中都可以发生,其成因较多,包括环境因子、植物本身的生理因子和化学因子[15-16]。研究结果表明,45%、65%、75%RH下Pn日变化呈“双峰”曲线,表现出明显的光合“午休”现象,85%RH呈“单峰”型曲线,未表现出“午休”现象,且净光合积累量随RH的增加不断增大。许大全[17]认为,增加空气相对湿度不仅能够消除植物叶片的“午睡”现象,而且显著地提高了净光合速率,本试验结果对此进行了进一步的证实。
引起光合速率降低的原因可以是气孔限制和非气孔限制。研究结果表明,不同RH处理下Ci的日变化趋势均呈“W”型曲线,与Pn的变化趋势相反,而Ls与Pn日变化趋势相似,即中午Pn下降时,Ci升高,而Ls降低,根据Farquhar et al.[18]对于光合“午休”现象的理论,可以推断出,槭叶草光合“午休”现象主要是叶肉细胞活性下降引起的非气孔限制,这与张宇[12]等的研究结果一致。
空气相对湿度与各光合参数的相关性分析表明,RH与Pn呈显著正相关,与Gs呈极显著正相关;而Pn与Ci呈显著正相关,与Ls呈显著负相关,说明Pn随RH的减小而降低的原因是气孔限制因素,这与王润元等[19]的研究结果一致,即空气相对湿度的增加促进了小麦在灌浆期和乳熟期叶片的气孔开放,使气孔导度增大,进而提高槭叶草叶片的净光合速率,反之,小麦的光合作用则受到限制。
凡是影响叶内外的蒸汽压差、气孔导度、叶边界层导度的各种生理和环境因子都会影响植物的蒸腾作用[20]。相关性分析表明,RH与Gs均呈显著正相关,说明增加空气湿度能够促进气孔开放;而Tr随RH增加而升高,可能是气孔导度增加导致的水气扩散阻力降低的原因所致。水分利用率是指植物消耗单位质量的水所固定的营养质量,是由净光合速率和蒸腾速率共同决定的[21];槭叶草WU。E由大到小依次为:75%、85%、65%、45%RH,说明较低的空气湿度不利于槭叶草对水分的利用。
综上所述,65% ~85%相对湿度均有利于槭叶草的生长和光合作用的增强。建议在园林绿化中将其配置在空气湿度较大的地方,如植被丰富的林缘、岸边绿化带或溪水旁。由于植物对水分的适应性不仅包括空气湿度的适应性,还包括土壤水分适应性,所以对槭叶草土壤水分的适应性还有待进一步研究,以期为其引种驯化和园林应用提供有力的理论依据。
[1] 乔恒,隋希英,陈建军,等.吉林省重点保护野生植物名录[J].吉林林业科技,2009:38(2):23-40.
[2] 王克凤,赵春莉,陈立飞.槭叶草离体快繁的研究[J].北方园艺,2012(5):135-136.
[3] 曲娴,刘建平,姜丽萍,等.槭叶草总黄酮对心衰鼠心功能的影响[J].北华大学学报:自然科学版,2000,1(4):306-313.
[4] 马全,杨淑娟,袁晓红.长白山早春植物与东北地区园林绿化[J].中国林副特产,2008(1):85-86.
[5] 宫敬利.槭叶草驯化栽培技术研究[D].北京:中国农业科学院,2007.
[6] 孙笑丛.对开蕨在干旱胁迫下形态生理及哈尔滨露地生境生长发育节律特征[D].哈尔滨:东北林业大学,2011.
[7] 丁爱萍,王瑞,张卓文,等.12 种园林植物耐荫性鉴定指标的筛选[J].植物生理学通讯,2009,45(1):55-59.
[8] 李合生.植物生理生化实验原理和技术[M].北京:高等教育出版社,2006:167-260.
[9] 梁宇,高玉葆,陈世苹.干旱胁迫下内生真菌感染对黑麦草实验种群光合、蒸腾和水分利用的影响[J].植物生态学报,2001,25(5):537-543.
[10] 尤伟忠,Hans K O K,房伟民,等.空气湿度对东方百合生长和切花品质的影响[J].园艺学报,2009,36(4):527-532.
[11] 许大全.光合作用效率[M].上海:上海科学技术出版社,2002.
[12] 张宇,宋敏丽,李利平,等.亚高温下不同空气湿度对番茄光合作用和物质积累的影响[J].生态学杂志,2012,31(2):342-347.
[13] 李金鹏,董然.3 种彩色叶玉簪光合日变化[J].东北林业大学学报,2011,39(10):56-70.
[14] 严海燕,董然,金光勋,等.遮阴对富贵草光合特性的影响[J].东北林业大学学报,2013,41(5):46-49.
[15] Osmond C B,Winter K,Powles S B.Adaptive significance of carbon dioxide cycling during photosynthesis in water-stressed plants[C]//Turner N C,Kramer P J (eds).Adaptation of plants to water and high temperature stress.New York:John Wiley & Sons,1980:139-154.
[16] 许大全.光合作用“午睡”现象的生态、生理与生化[J].植物生理学通讯,1990(6):5-10.
[17] 许大全.田间小麦叶片光合作用“午睡”现象的研究[J].植物生理学报,1984,10(3):269-276.
[18] Farquhar G D,Sharkey T D.Stomatal conductance and photosynthesis[J].Ann Rev Plant Physiol,1982,33:317-345.
[19] 王润元,杨兴国,赵鸿.半干旱雨养区小麦叶片光合生理生态特征及其对环境的响应[J].生态学杂志,2006,25(10):1161-1166.
[20] 于贵瑞,王建林.植物光合、蒸腾与水分利用的生理生态学[M].北京:科学出版社,2010.
[21] 赵国禹,董然,赫微微.复序橐吾的光合特性及其影响因子[J].东北林业大学学报,2011,39(1):41-43.
猜你喜欢
现代农业科技(2022年10期)2022-12-17
花卉(2021年1期)2021-01-11
新课程·下旬(2017年7期)2017-08-14
Coco薇(2016年5期)2016-06-03
考试周刊(2016年6期)2016-03-11
少儿科学周刊·儿童版(2015年10期)2015-11-07
西北林学院学报(2015年5期)2015-01-03
植物营养与肥料学报(2011年2期)2011-10-26
植物营养与肥料学报(2010年3期)2010-11-16
百科知识(2009年24期)2009-07-09
