
原料乳中高产蛋白酶嗜冷菌的分离及产酶培养基的研究
2014-03-03 08:27张阳旸陈万义王渊龙
食品工业科技 2014年20期
张阳旸,刘 景,陈万义,王渊龙,任 婧,*
(1.乳业生物技术国家重点实验室,光明乳业股份有限公司研究院,上海 200436;2.上海海洋大学食品学院,上海 201306)
原料乳中高产蛋白酶嗜冷菌的分离及产酶培养基的研究
张阳旸1,2,刘 景1,陈万义1,王渊龙1,任 婧1,*
(1.乳业生物技术国家重点实验室,光明乳业股份有限公司研究院,上海 200436;2.上海海洋大学食品学院,上海 201306)
以原料乳为对象,使用脱脂乳平板筛出三株有较强蛋白酶水解性的嗜冷菌,通过时间-酶活曲线复筛出一株高产蛋白酶的嗜冷菌Px-1。针对其产蛋白酶培养基进行了正交优化,研究了其产酶培养基的主要成分,结果表明酵母浸出粉对产酶影响显著。通过本实验最终确定了最佳培养基组成:蔗糖4%,α-乳糖4%,葡萄糖2%;酵母浸出粉0.7%,干酪素0.5%,胰蛋白胨0.3%以及0.05%的吐温80。在此条件下,Px-1产蛋白酶活为42.5451U/mL,高于优化前的测定值30.2291U/mL。
原料乳,嗜冷菌,蛋白酶,培养基,优化
牛奶中的嗜冷菌几乎覆盖了牛奶中常见的细菌的所有类群[1-2],由于嗜冷菌对牛乳的污染造成了大量乳制品的腐败[3]。目前,对于牛乳中的嗜冷菌,已经有一些先进的快速检测方法,如直接荧光过滤法、电阻抗法、PCR法、流动血细胞法及酶联免疫吸附技术等[4-9],但这些方法成本过高。为了解决这个问题,仅从嗜冷菌菌体本身考虑已经无法达到快速检测的目的,因此可以从其分泌的蛋白酶入手,通过对其酶活性的检测以及酶活和菌数之间建立的联系达到快速检测原料乳中嗜冷菌数的目的,此前已有研究表明,牛乳中的嗜冷菌数和蛋白酶活性存在一定的关系[10]。这为建立嗜冷菌快速检测方法提供理论依据。
但由于原料乳中嗜冷菌分泌的蛋白酶含量很低[11],自 然 其 活 力 也 不 高 ,因 而 限 制 了 检 测 的 精 确度。而培养基的成分是影响嗜冷菌发酵水平的关键因素,因此,如何提高产酶培养基灵敏度,即如何优化产酶培养基便是本研究所要讨论的问题。
1 材料与方法
1.1 材料与仪器
乳样 取自上海地区牧场;福林试剂、三氯乙酸、碳酸钠、磷酸二氢钠、磷酸氢二钠、氢氧化钠、酪氨酸、胰蛋白胨、酵母浸膏、葡萄糖、脱脂奶粉、琼脂粉、α-乳糖、蔗糖、淀粉、麦芽糖、大豆蛋白胨、硫酸铵、尿素、干酪素、吐温20、吐温60、吐温80、SDS 均为分析纯,购于国药集团化学试剂有限公司。
MLS-3750高压灭菌锅 日本SANYO;Minispinplus台式离心机 德国艾本得eppendorf;BPH-9082恒温培养箱 上海一恒科技有限公司;SpectraMax M5酶标仪 美国Molecular Devices公司。
1.2 实验方法
1.2.1 培养基 MPC琼脂培养基(1000mL):胰蛋白胨5g,酵母浸膏2.5g,葡萄糖1.0g,脱脂奶粉1.0g,琼脂15g,蒸馏水,pH调节至7.0±0.1。
种子培养基(1000mL):胰蛋白胨10g,酵母浸膏5g,氯化钠10g,琼脂15g,蒸馏水,pH调节至7.0±0.1。
脱脂乳平板(1000mL):脱脂乳粉100g,琼脂粉15g,pH调节至7.0±0.1。
产酶基础培养基(1000mL):葡萄糖20g,酵母膏10g,KH2PO42.0g,MgSO4·7H2O 0.5g,CaCl20.2g,pH调节至6.5。
1.2.2 菌株筛选
1.2.2.1 初筛 将原料乳制备成不同稀释浓度的菌悬液,选择3个稀释度,每个稀释度取0.1mL涂布于脱脂乳平板,7℃条件下培养10d。将脱脂乳平板上能形成透明圈的菌落,挑出在脱脂乳平板上纯化分离2~3次,分离纯化出的单菌落点接于脱脂乳平板培养基上,7℃培养10d,测量并计算水解圈与菌落直径比值HC[12]。
1.2.2.2 复筛 将初筛中透明圈直径与菌落之比较大的菌株接种于种子培养基中,180r/min摇瓶培养10h,按5%(v/v)的接种量转入复筛培养基中,离心5min,取上清液,利用福林-酚法测定菌株产蛋白酶的粗酶酶活[13],通过时间-酶活曲线筛选出产酶酶活最高的菌株[14]。
1.2.3 单因素实验
1.2.3.1 不同碳源对产酶的影响 分别选择葡萄糖、蔗糖、麦芽糖、α-乳糖、淀粉作为碳源,以酵母膏作为氮源,其他成分同产酶基础培养基,单因素实验选择合适的碳源。
1.2.3.2 不同氮源对产酶的影响 选择酵母膏、大豆蛋白胨、胰蛋白胨、硫酸铵、尿素、硝酸钠、酪蛋白作为氮源,以葡萄糖作为碳源,其他同产酶基础培养基,单因素实验选择合适的氮源。
1.2.3.3 不 同 表 面 活 性 剂 对 产 酶 的 影 响 选 择 吐温-20、吐温-60、吐温-80,SDS作为表面活性剂,其他同产酶基础培养基,单因素实验选择合适的表面活性剂。
1.2.4 发酵培养基的正交设计 通过单因素实验,筛选出蔗糖、α-乳糖、葡萄糖、酵母浸出粉、干酪素、胰蛋白胨、吐温80为影响因子,在单因素实验的基础上,每个因子设定三个水平,正交设计出18组实验(表1),以产蛋白酶活力为标准进行优化组合筛选。
2 结果与分析
2.1 菌株筛选结果
2.1.1 菌株初筛结果 从图1和表2可知,实验筛选出了9株能产生明显的透明圈的嗜冷菌,通过比较不同的菌株在脱脂乳平板上的透明圈/菌落直径比(HC值),可以看出,菌株Px-2,Px-4,Px-6,Px-7,Px-8,Px-9的HC值均在2.00以下;而菌株Px-1,Px-3,Px-5的HC值均在2.4以上,由此初步判定,菌株Px-1,Px-3,Px-5相对另外6株菌株具有较强的产蛋白酶能力,因此选用这三株菌进行复筛以确定其中高产蛋白酶的嗜冷菌。

表1 产酶培养基正交组合实验设计(L1873)Table 1 Orthogonal design of enzyme producing medium(L1873)
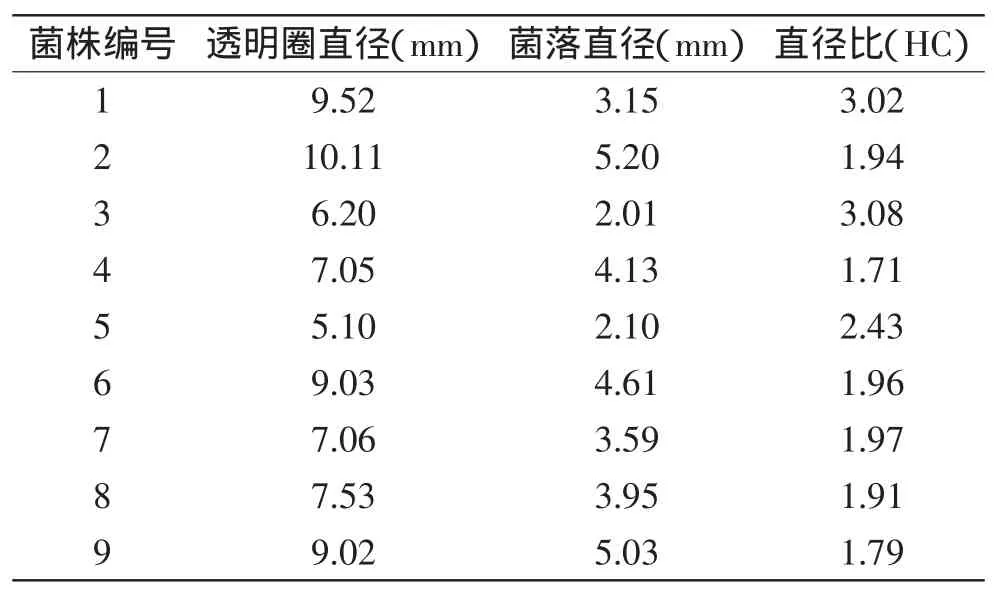
表2 产蛋白酶嗜冷菌透明圈/菌落直径比Table 2 Transparent circle diameter-to-colony diameter ratios of 9 bacterial isolates
2.1.2 菌株复筛 图2为蛋白酶活力标准曲线,由图2可以看出,在波长范围内660nm处的吸光光度值与蛋白酶活性有良好的线性关系,检测限量为0.995。因此可以确定,采用此法检验嗜冷菌产蛋白酶酶活,方法可靠。对初筛的3株菌株Px-1,Px-3,Px-5进行分离纯化后,在产酶基础培养基中摇瓶发酵48h,温度28℃,转速为180r/min,每6h取样测定三株菌的产蛋白酶酶活,绘制时间-酶活曲线图,结果见图3。
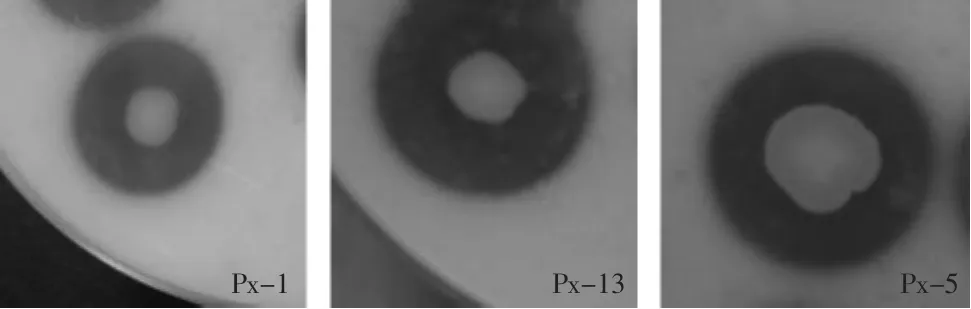
图1 Px-1,Px-3,Px-5产生的水解圈Fig.1 Hydrolytic ring of Px-1,Px-3,Px-5
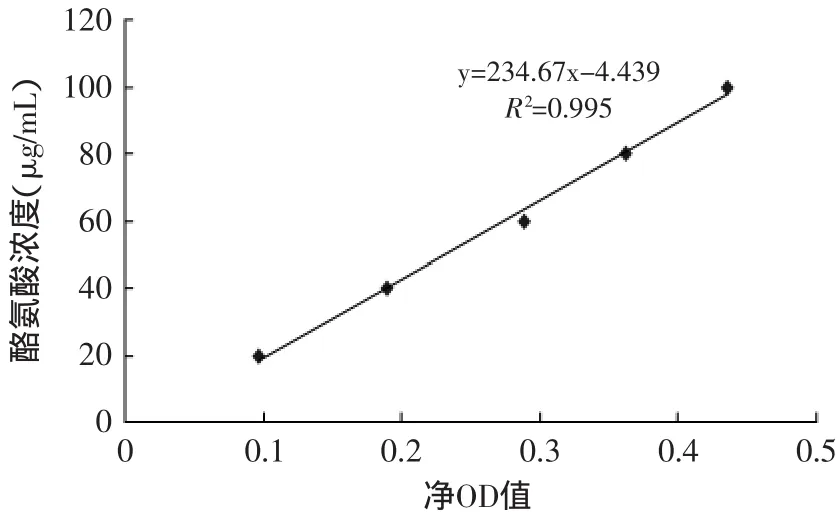
图2 蛋白酶活性标准曲线Fig.2 The standard curve of protease activity

图3 Px-1,Px-3,Px-5的蛋白酶酶活-时间曲线Fig.3 The enzyme-time activity curve of Px-1,Px-3,Px-5
由图3可知,随着发酵时间的延长,Px-1,Px-3,Px-5的产酶活性逐渐达到最高点,之后开始下降。其中Px-1在培养时间0~30h内酶活不断增加,并在30h达到最高,为30.3488U/mL。30h后酶活开始逐渐降低;Px-3在30h酶活达到最高为24.5566U/mL;Px-5在24h酶活达到最高为7.1289U/mL。由此可以推出,发酵培养时间可能是影响酶活的重要因素,菌液接入发酵液后短时间内酶活力较低,可能是因为此时菌数不多,且大多数菌还处于生长调整期,主要进行的是菌体生长活动。而随着发酵时间逐渐增加,菌体数也会随之显著增加,到达稳定期后,细菌数量保持稳定,开始分泌代谢产物,因此在这段时间内,酶活随着发酵时间的增加逐渐增加。在稳定期后,随着培养时间的增加,细菌开始进入衰亡期,菌体老化,生物量下降,因此培养时间过长反而出现酶活降低的情况[15]。因此本实验通过比较三株菌在相对最佳酶活时间点处的酶活,以在30h处酶活最高的Px-1作为研究对象。
2.2 产酶培养基的优化
2.2.1 不同碳源对蛋白酶活力的影响 不同碳源对蛋白酶活力的影响如图4所示,Px-1在5种不同碳源条件下培养时,碳源对蛋白酶的活力具有一定的影响,影响力大小依次为蔗糖>α-乳糖>葡萄糖>麦芽糖>淀粉。相对其他碳源来说,蔗糖对嗜冷菌具有很好的增殖菌体作用[16]。
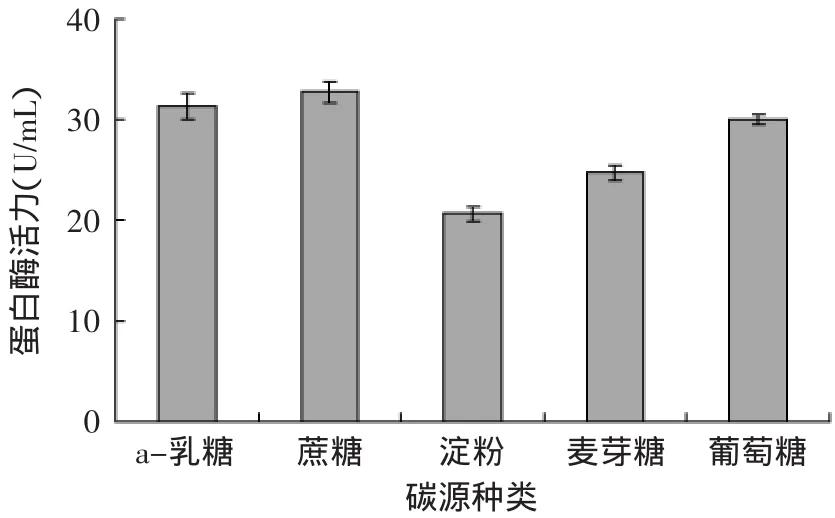
图4 不同碳源对Px-1产蛋白酶活力的影响Fig.4 Effect of different carbon sources on enzyme activity of protease
2.2.2 不同氮源对蛋白酶活力的影响 不同氮源对蛋白酶活力的影响如图5所示,Px-1在5种氮源条件下培养时,氮源对蛋白酶活力的影响较为显著,其中酵母浸出粉影响最大,酶活力达35.2767U/mL、其次分别是干酪素、胰蛋白胨、尿素和大豆蛋白胨。
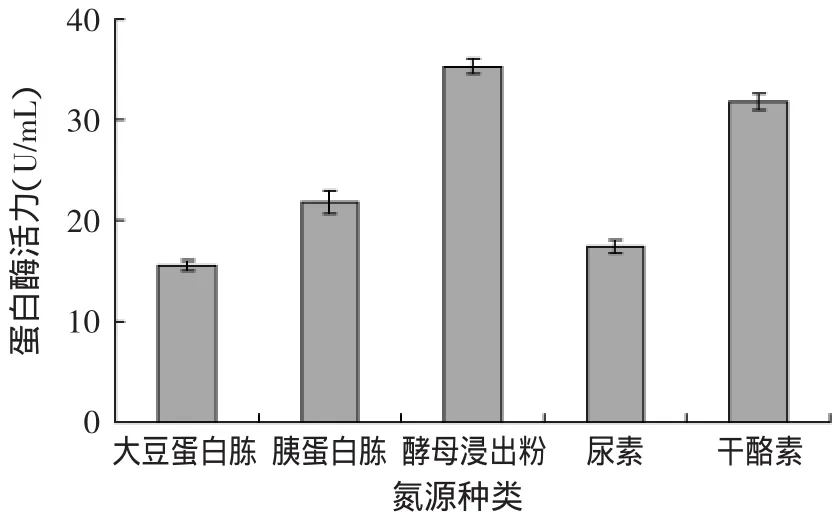
图5 不同氮源对Px-1产蛋白酶活力的影响Fig.5 Effect of different nitrogen sources on enzyme activity of protease
2.2.3 不同表面活性剂对蛋白酶活力的影响 由图6可知,表面活性剂对Px-1产酶的影响一次为吐温80>吐温60>SDS>吐温20。吐温80、吐温60显著的提高了蛋白酶的活力,可能是因为表面活性剂可以与细胞膜相互作用,增加细胞的通透性,使更多的酶从细胞内透过细胞膜泄漏出来,从而有利于胞外酶的分泌,从而提高了酶的产量[17]。
2.2.4 正交实验结果 图7为嗜冷菌Px-1在不同培养基中的发酵产生的蛋白酶酶活曲线,对正交设计的18个配方进行发酵产酶实验,结果表明,9号配方产蛋白酶活力最强,为42.5510U/mL。配方为蔗糖4%,α-乳糖4%,葡萄糖2%;酵母浸出粉0.7%,干酪素0.5%,胰蛋白胨0.3%以及体积分数0.05%的吐温80。在所有的实验组中,除了1、5、11、12、14、15、18组以外,其他配方组产蛋白酶的活力均在30.3488U/mL以上,均高于优化前Px-1在30h时的产蛋白酶酶活力。
2.2.5 正交实验结果方差分析 各因素对Px-1产蛋白酶的影响程度可以由p值确定,利用SPSS软件分析实验结果[18],得出各因素的p值,结果如表3所示。由于p值越小越显著,结果表明,影响Px-1产酶的最显著因素是酵母浸出粉,其p值为0.011<0.05。其余因素对产酶影响并不显著。
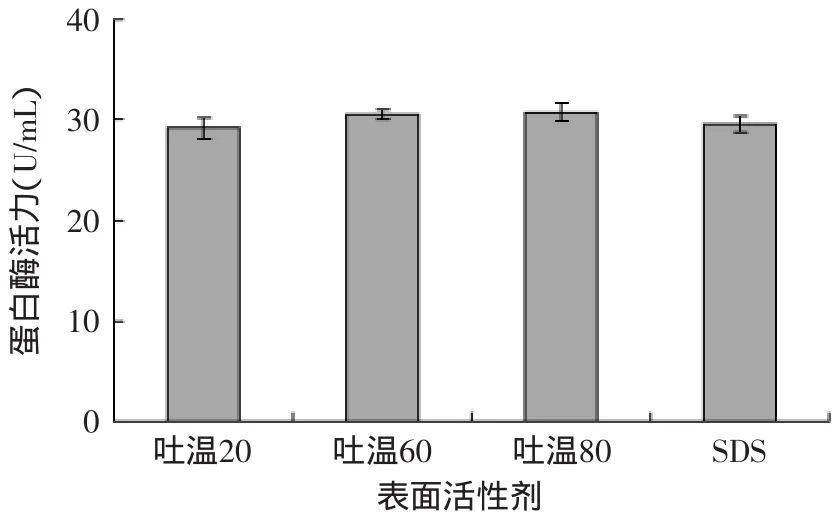
图6 不同表面活性剂对Px-1产蛋白酶活力的影响Fig.6 Effect of different surfactants on enzyme activity of protease
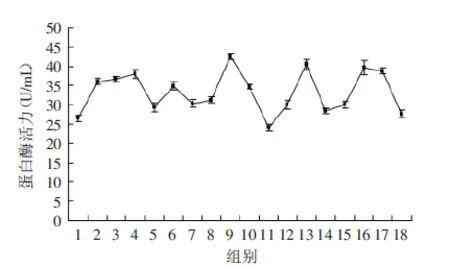
图7 Px-1在优化后产酶培养基中的酶活曲线Fig.7 The enzyme activity curve of Px-1 in optimized medium

表3 正交实验结果方差分析Table 3 Variance analysis of orthogonal experiment results
3 结论
本实验从原料乳中筛选到三株显著产蛋白酶嗜冷菌Px-1、Px-3、Px-5,通过时间-产酶曲线确定其中高产蛋白酶的菌株Px-1,产酶培养的最佳时间为30h,初始产酶水平为30.3488U/mL。通过单因素实验和正交确定了最佳产蛋白酶培养基组成:蔗糖4%,α-乳糖4%,葡萄糖2%,酵母浸出粉0.7%,干酪素0.5%,胰蛋白胨0.3%以及体积分数0.05%的吐温80;通过对正交实验结果的方差分析可知,酵母浸出粉对Px-1产酶影响显著,在优化后的产酶培养基中培养30h后所测得的最高蛋白酶活力高于基础产酶培养基中培养30h后的蛋白酶活力,因而通过在优化产酶培养基上的培养,可以提高检测的精确度。
[1]T.Sørhaug.Psychrotrophs and their enzymes in milk and dairy products:Quality aspects[J].Trends in Food Science& Technology,1999(8):35-41.
[2]Samagh,Cunningham.Numerical Taxonomy of the Genus Pseudomonas from Milk and Milk Products[J].Journal of Dairy Science,1971(55):19-24.
[3]韩中惠,黄艾祥,毛华明,等.嗜冷菌对牛乳的危害及检测控制[J]. 中国奶牛,2012(19):42-44.
[4]Pettipher.Review:The direct epifluorescent filter technique [J].Journal of Food Technology,1986,21(5):535-546.
[5]周向华,王衔彬,叶兴乾.电阻抗法在食品微生物中快速检测中的应用[J]. 粮油加工与食品机械,2003(10):73-75.
[6]Gutierrez R,Garcia T,Gonzalez I.A quantitative PCR-ELISA for the rapid enumeration of bacteria in refrigerated raw milk[J]. Journal of Applied Microbiology,1997,83(4):518-523。
[7]N Yamaguchi.H Ohba,M Nasu.Simple detection of small amounts of Pseudomonas cells in milk by using a microfluidic device[J].Letters in Applied Microbiology,2006,43(6):631-636.
[8]R Gutiérrez,I González,T García.Monoclonal antibodies and an indirect ELISA for detection of psychrotrophic bacteria in refrigerated milk[J].Journal of Food Protection,1997,60(1):23-7. [9]庞伟华.牛乳中嗜冷菌检测方法的研究进展[J]. 中国乳业,2013(1):58-60.
[10]王辉,吕加平,迟玉杰.牛乳中嗜冷菌数与胞外蛋白酶活性相关性的研究[J]. 食品工业科技,2008,29(2):84-86.
[11]张书文,刘鹭,李红娟,等.荧光假单胞菌胞外蛋白酶的纯化及其特性研究[J]. 食品科学,2012,33(9):206-210.
[12]庞宗文,李敏,李树波,等.产蛋白酶毛霉的分离筛选及发酵豆粕产大豆肽的初步研究[J]. 现代食品科技,2010(9):956-961.
[13]顾瑾麟,夏永军,张红发,等.开菲尔粒中高产蛋白酶菌株的筛选及培养基优化[J]. 现代食品科技,2013,29(3):558-562.
[14]廉立慧,高丽君,王德才,等.高温蛋白酶产生菌的筛选及其产酶条件和酶学性质分析[J]. 生物技术通报,2011(3):175-179.
[15]任静,宋兴舜,张兰威.原料乳中典型嗜冷菌增菌培养条件优化[J]. 中国乳品工业,2009,37(6):13-15.
[16]刘坚真,黄珂,陈伟力.培养基对细菌总数测定的影响及其改进[J]. 食品科学,2003(6):105-108.
[17]薛林贵,景春娥,赵旭,等.重离子诱变技术选育碱性蛋白酶高产菌株[J].微生物学通报,2010,37(6):845-851.
[18]张秋会,李苗云,赵改名,等.牛乳酪蛋白水解特性研究[J].现代食品科技,2010,26(10):1130-1133.
Study on screening and optimization of fermentation media of high protease-producing psychrotrophic bacteria
ZHANG Yang-chang1,2,LIU Jing1,CHEN Wan-yi1,WANG Yuan-long1,REN Jing1,*
(1.State Key Laboratory of Biotechnology Industry,Bright Dairy Research Institute,Shanghai 200436,China;2.College of Food of Shanghai Ocean University,Shanghai 201306,China)
Three psychrotrophic strains coded Px-1,Px-3 and Px-5 which could produce protease and have obvious hydrolysis circles on defatted milk powder plate,that were isolated from raw milk.The stains were isolated again by time-enzyme activity curve.The strain Px-1 had the strongest protease activity.Composition of fermentation medium was optimized by quadratic design.The optimum fermentation medium composition was as following:4%sucrose,4%alpha lactose,2%glucose,0.7%yeast extract powder,0.5%casein,0.3%tryptone and 0.05%tween 80.Under the optimum conditions ,the protease activity of Px-1 could reach 42.5451U/mL ,higher than the value before optimization 30.2291U/mL.
raw milk;psychrotrophic bacteria;protease;fermentation medium;optimization
TS252.1
A
1002-0306(2014)20-0204-04
10.13386/j.issn1002-0306.2014.20.036
2013-12-17
张阳旸(1989-),女,硕士研究生,研究方向:食品微生物学与分子生物学。
* 通讯作者:任婧(1980-),女,高级工程师,研究方向:乳酸菌分子生物学。
国家“十二五”科技支撑计划课题(2012BAD12B08);国家“973”计划课题(2012CB723706)。
猜你喜欢
小学生学习指导(爆笑校园)(2020年3期)2020-06-05
疯狂英语·初中天地(2018年6期)2018-11-24
山东工业技术(2016年15期)2016-12-01
阅读(中年级)(2016年9期)2016-05-14
中国酿造(2016年12期)2016-03-01
中国酿造(2016年12期)2016-03-01
应用海洋学学报(2015年3期)2015-11-22
中国酿造(2014年9期)2014-03-11
食品工业科技(2014年9期)2014-03-11
中国食用菌(2013年2期)2013-11-21
