
外来植物大花老鸭嘴在迁入地的传粉机制和繁殖策略
2014-02-10 00:43唐光大成建飞谢振兴陈瑾林翠菊
生态环境学报 2014年6期
唐光大,成建飞,谢振兴,陈瑾,林翠菊
华南农业大学林学院,广州 510640
外来植物大花老鸭嘴在迁入地的传粉机制和繁殖策略
唐光大*,成建飞,谢振兴,陈瑾,林翠菊
华南农业大学林学院,广州 510640
外来植物在迁入地的生存和繁殖策略,是入侵生物学研究的热门话题之一。开展入侵植物的繁殖策略研究,有助于深入了解入侵生物在迁入地的生存繁衍机制,对入侵植物在可控范围内的适当使用具有重要的指导意义。通过野外传粉昆虫观测、光学和电镜扫描观测和人工控制授粉等方法,对外来植物大花老鸦嘴(Thunbergia grandiflora)在广州和深圳的4个居群的开花物候、花部结构、花粉活力、传粉昆虫及其行为等传粉生物学特征进行了研究。结果表明:大花老鸦嘴的单花花期为1 d,上午10:00时花粉活力最高,长雄蕊花粉活力可达73.92%,短雄蕊花粉活力可达71.54%,随后花粉活力逐渐下降。主要传粉者为扁柄木蜂(Xylocopa latipes),开花当天访花高峰期的9:00—10:00,单花访花频率可达5 h-1以上,之后开始逐渐减少,14:00之后未观测到扁柄木蜂访花,18:00后仅剩蚂蚁等体型较小的昆虫长期栖息于花序或花蕾。控制授粉实验结果显示,无论是人工异株授粉、自花授粉,还是自然授粉,结果率均为0,4个居群均未见结实。光学显微镜下子房发育完整;电镜扫描观测结果显示其花粉在柱头上能萌发,但花粉管不能延伸到子房位置。目前在园林栽培的大花老鸭嘴均以块根或茎为繁殖体培育苗木,虽然这些植株均能开出鲜艳的花朵,花粉传递者也充足,但未见任何结果现象。因此,初步推测本次研究区域内的大花老鸭嘴种群可能来自相同或相近的无性系,故不同居群间异株授粉的不结果现象可能为“自交不亲和”,或“染色体多倍化”所致。
大花老鸦嘴;传粉生物学;扁柄木蜂;自交不亲和
由于人类有意或无意的活动,将植物种类带到其自然演化区域以外,这些被外力驱动迁移到新领地的植物,称为外来植物(alien plants) (Richardson等, 2000)。部分外来植物会形成入侵和造成危害(Mack等, 2000),其入侵性往往是一系列功能性状和环境因子综合作用的结果(Abhilasha和Joshi, 2009; Dawson等, 2009)。许多植物通过有性繁殖,产生大量质量轻、容易扩散的种子,在新分布区域迅速传播,形成大面积的单一群落,危害农田和湿地生态系统。尤其是菊科中的外来植物,如三叶鬼针草(Bidens pilosa) (洪岚等, 2004)、假臭草(Eupatorium catarium) (钟军第等, 2014)、飞机草(Chromolaena odorata) (柴伟玲等, 2014)、紫茎泽兰(Ageratina adenophora) (姜勇等, 2013)等。但也有部分植物通过无性繁殖,扩大分布区域,如南美蟛蜞菊(Wedelia trilobata) (杨东娟和朱慧, 2008)、爵床科(Acanthaceae)的部分植物(Meyer, 2004)。植物的入侵过程也是该植物入侵个体适应新生境的过程,往往伴随着生长和繁殖方面快速的适应性变化,因此,植物入侵现象也为研究植物繁育系统的进化提供了绝佳机会(Lee, 2002)。由于繁育系统不仅在入侵过程中发生了适应性变化,也反过来对植物入侵机制、途径等有着深刻的影响,因此,研究入侵植物的繁育系统变化不但可以了解植物繁育系统的进化模式,也是生物入侵研究和管理实践中的核心问题之一(任明迅等, 2004)。另外,研究外来植物的繁殖策略,对有效控制其分布范围,并加以适当利用,具有一定的理论指导意义。
大花老鸦嘴 (Thunbergia grandiflora)为爵床科山牵牛属的常绿藤本植物,又名大花山牵牛、山牵牛、大邓伯花。喜温暖湿润及阳光充足的环境,不耐寒,稍耐阴,在排水良好、湿润的沙质壤土上生长良好。适宜温度为15~30 ℃。以茎缠绕进行攀援,长势快,分枝能力甚强,病虫害少。原产印度北部和孟加拉国,现广泛种植于中国华南、东南亚和澳大利亚、新西兰等地。我国台湾早在1910年就有引种记录,目前在华南地区普遍栽培(黄芳, 2009),主要运用于藤架绿化、庭园造景。近年来,随着立体绿化的快速发展,大花老鸦嘴的快速生长和覆盖能力,加之淡紫色的花大而艳丽,观赏价值高,在城市园林绿化中,多用于攀援棚架、阳台、天台及城市立交等垂直绿化方面。然而,大花老鸦嘴是一种具有较强入侵性的外来物种,在澳大利亚东海岸、夏威夷、新加坡等地已经造成较严重的危害(Dawson等, 2008)。
目前,国内对大花老雅嘴的研究,主要集中在园林应用中的耐荫性(陈国菊等, 2001)、组织培养(陆耀东等, 2008)、扦插繁殖(吴冲和史正军, 2008)、和部分药用价值(江纪武, 2003)等方面。有关该物种在迁入地的繁殖特性和策略方面,尚未见相关研究报道。本文报道了大花老鸭嘴在广州和深圳的开花特性和传粉机制,较详细地探讨了大花老鸭嘴在华南地区只开花不结果的可能原因,并推测大花老鸭嘴在本次研究区域内主要借助人为力量进行无性繁殖,实现其迅速抢占生境,生存繁衍的目的。
1 材料与方法
1.1 研究地概况
广州地处亚热带北缘,属海洋性亚热带季风气候,温暖多雨、光热充足、夏季长、霜期短。全年平均气温为20~22 ℃,7月最热,月平均气温达28.7 ℃,1月最冷,月平均气温为9~16 ℃,平均相对湿度77%。深圳位于广东省南部,珠江口东岸,也属于亚热带海洋性气候。年平均气温22.4 ℃。雨量充足;7月平均气温最高,为28.9 ℃,1月平均气温最低,为15.4 ℃,相对湿度也较高。
1.2 花形态及物候观测
2008—2010年持续3年的7月—8月在广州华南农业大学和长湴小学、深圳市园林科学研究所、深圳市兰科植物保护研究中心,各随机选取约20朵刚开放的花朵,测量其花冠直径、花冠长度、雄蕊长度、雌蕊长度、柱头直径、柱头与花药之间的距离,并描述雄蕊上的毛被特征等;并于2009年在广州华南农业大学校内采集大花老鸭嘴的花样品,进行光学显微镜(Optical microscope)观测和电镜扫描(Scanning electron microscope, SEM)观测。
1.3 繁育系统检测
分别在2008—2010年在广州和深圳各选取2个大花老鸦嘴居群,在花蕾期套袋,花刚开放时,对新鲜花开展以下人工控制授粉实验:1)自然授粉;2)去雄套袋不授粉;3)去雄不套袋自然授粉;4)人工异株花粉;5)人工自花花粉;6)不去雄,套袋,自然授粉。待花冠萎蔫,花粉和柱头失去活力之后,解开袋子,减小套袋对子房发育的影响。30天后检测和统计各处理的结实率。
1.4 花粉活力和柱头可授性测定
花开放后取新鲜花粉,置于载玻片上,滴加含0.5%TTC(2, 3, 5-三苯基氯化四氮唑)的蔗糖溶液,放入内有湿滤纸的培养皿中,在37 ℃下的黑暗环境下放置,每隔2 h在显微镜下统计花粉颜色变红和花粉管萌发的数量,并计算出有活力的花粉所占的比例。柱头可授性测定参照彭东辉等(2008)的方法,用在不同花期对柱头进行人工授粉,观测其结实率来估测。
1.5 昆虫传粉行为观测与昆虫传粉效率的测定
随机标记30个即将开放的花蕾,从开花当天8:00开始,持续对其访花者进行观察,直至花冠枯萎。记录访花昆虫的种类、访问次数、访花时间、携带花粉的情况,同时描述昆虫的传粉行为。
2 结果分析
2.1 花形态结构及物候观测
大花老鸦嘴单花花期为1 d,在广州和深圳的4个居群内全年均可开花(图1A、B)。早上8:00左右花冠裂片全部展开,4枚雄蕊的花药均开裂,散播花粉(图1B)。花部观察和测量结果见表1,花盛开时花冠直径7.5~9.1 cm,花冠筒深度3.0~3.8 cm。花冠筒上部白色,底部浅黄色,花冠裂片5,覆瓦状排列,蓝紫色或淡白色(图1B);花开放当天下午13:00以后,花冠颜色开始变淡,逐渐萎蔫;雄蕊4枚,分为前后2排,并列,后2枚略短(图1C、D),花药内藏,纵裂(图1E),长雄蕊的药囊边缘芒状附属物呈鬃毛状,浓密,短雄蕊的芒状附属物较稀疏(图1C、D);雌蕊长于雄蕊,成熟柱头乳白色,漏斗状,高于花药,长雄蕊的花药与柱头间距约0.25 cm;柱头的近轴裂片波状折叠,远轴裂片弯曲成铲状(图1F);子房基部与花冠筒粘结部位分泌的花蜜清晰可见(图1H)。光学显微镜下,大花老鸭嘴的子房发育正常,未见子房败育现象(图1G)。
电镜观测显示,大花老鸭嘴的花粉粒呈圆球形,直径约62 μm,表面具螺旋状萌发孔,外壁纹饰略呈颗粒状(图2A),柱头边缘略波浪状,在一侧皱褶,形成略似铲状的结构,柱头表面具密集突起的短毛(图2B、C)。
2.2 繁育系统检测
在2008—2010年的3次人工控制授粉试验中,无论是人工自花受粉、异株授粉,还是标记的自然授粉,所有处理的结实率均为0。而且在4个居群内的所有植株上,均未观测到大花老鸭嘴结果的现象。除了选作研究对象的4个居群,我们还观测了广州华南植物园内、深圳市龙岗区大鹏半岛等地分布的大花老鸭嘴居群,均未发现大花老鸭嘴的自然结果现象。对人工异株授粉后的柱头和花粉粒电镜扫描观测发现,花粉粒在柱头上能够萌发和生长出花粉管,但仅长出约70 μm左右(图2D),未见花粉管伸长,延伸至子房的现象。

图1 大花老鸭嘴居群、花结构和传粉昆虫Fig. 1 The polulation, flower and pollinator of Thunbergia grandiflora

表1 大花老鸦嘴的花部结构特征Table 1 Flower structure of Thunbergia grandiflora cm
2.3 花粉活力及柱头可授性
花粉活力测定过程中,将大花老鸦嘴的花药分成长雄蕊和短雄蕊2组,分别测定。在开花当天,长雄蕊和短雄蕊的花粉活力均呈逐渐下降的变化趋势,二者的花粉活力差异较小。上午10:00时花粉活力最高,长雄蕊花粉活力可达73.92%,短雄蕊花粉活力可达71.54%,随后花粉活力逐渐下降(图3)。开花第2天凌晨,花冠与雄蕊一起脱落。
使用不同花期人工异株授粉的方式,检测其结实率,估测其柱头可授性。人工异株授粉的结实率结果显示,所有时间段的异株授粉的花,均未结果,所有处理的结实率均为0,未能体现出大花老鸭嘴的柱头在不同花期的可授性差异。
2.4 昆虫传粉行为
在4个居群内所观测到的大花老鸦嘴的访花昆虫主要有扁柄木蜂(Xylocopa latipes)和蚂蚁(Ants),而只有扁柄木蜂是其有效的传粉者,未见蚂蚁携带花粉。扁柄木蜂的体型与花冠筒的大小相近,在传粉者取食花蜜过程中,其头部前半部分与雄蕊基部紧密接触,雄蕊上的鬃毛与传粉者前胸背部和触角接触,在相互摩擦过程中,纵裂的花药囊里的花粉散落于传粉者背部(图1J、K)。当传粉者取食完毕,从花冠筒退出时,前脚往回收,头部向上抬起,整个背部均与花药紧密接触,花粉再次大量散落于其背部(图1I、K),当传粉者飞到其它植株的花上采花蜜时,带走的花粉将会粘到柱头上,完成异花授粉过程(图1I)。
在大花老鸦嘴的开花季节,整个花序上持续有许多蚂蚁在花序轴、花冠筒花丝和花柱部位逗留,并爬至花冠筒基部取食花蜜(图1L)。蚂蚁来回穿梭于花冠筒、花丝和花柱等部位,但未观测到蚂蚁身上携带大花老鸭嘴的花粉,而且绝大多数蚂蚁仅在同一植株内的不同花序上活动,所以蚂蚁仅仅可能造成极少量的自花授粉。

图2 大花老鸭嘴的花粉和柱头的电镜扫描观测Fig. 2 Pollen and stigma of T. grandiflora under SEM
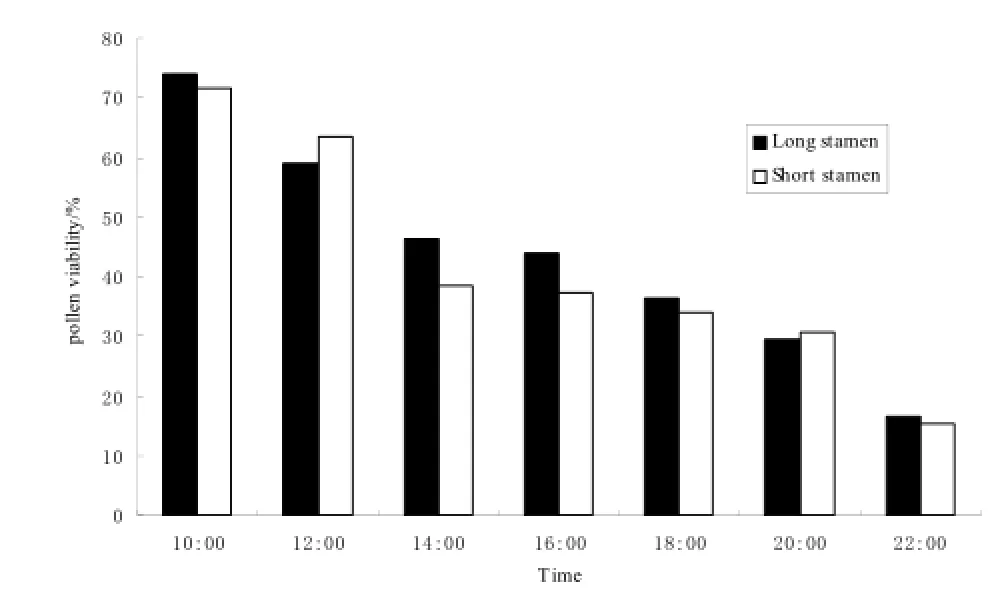
图3 大花老雅嘴长短雄蕊的花粉活力Fig. 3 The pollen viability of long and short stamens of T. grandiflora
2.5 昆虫访花频率与时间
大花老鸦嘴的单花花期为1 d,有效传粉者扁柄木蜂的访花频率随着时间呈先上升后下降的变化趋势,扁柄木蜂的访花高峰期为开花当天的9:00—10:00,单花访花频率可达5 h-1以上,之后开始逐渐减少,到13:00—14:00时,访花者明显减少,14:00之后未观测到扁柄木蜂访花 (图4)。
开花当天8:00—11:00期间,扁柄木蜂在花冠上的平均停留时间呈下降趋势,由8:00—9:00约10 s逐渐下降到9:00—10:00约5.4 s;在10:00—12:00期间,昆虫在花冠上的停留时间变化较小,维持在5.6 s左右;在12:00—13:00的时间里,昆虫在花上的平均停留时间略有上升,达到6.4 s,访花昆虫减少;13:00—14:00期间,扁柄木蜂的访花时间快速缩短(图5)。14:00之后,当天开放的花开始逐渐萎蔫,花冠皱缩,枯萎,无传粉昆虫访花。
2.6 花部构成对昆虫吸引力的检测
通过人工切除花冠,观测传粉者的访花频率,检测花冠对传粉者的吸引力。实验结果如图5所示,完全切除花冠的花,整个开花期间均无传粉者访花;横向和纵向切除一半花冠的花,传粉者的访花频率相近,约为2~2.8 n·h-1,早上8:00—10:00访花频率最高,随后逐渐下降;不切除花冠的传粉者访花频率最高,整个花期内的访花频率为2.3 n·h-1,最高可达3.8 n·h-1,说明淡紫色花冠对传粉者扁柄木蜂具有较强的视觉吸引和定位作用(图6)。

图4 传粉者扁柄木蜂的访花频率Fig. 4 The visiting flowers’ frequency of Xylocopa latipes
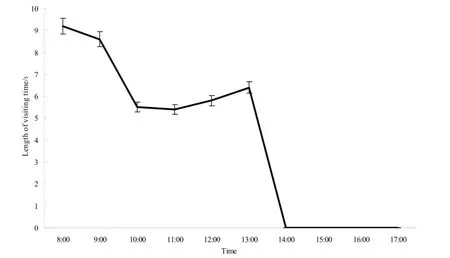
图5 扁柄木蜂的访花时间Fig. 5 The visiting time of X. latipes
3 讨论
3.1 大花老鸭嘴的繁殖方式
外来植物在迁入地采取的与环境相适应的繁殖策略和快速繁殖能力,是其能否在新的生境中快速扩张的关键因素之一。繁殖能力的大小对外来植物能否成功定居和扩散具有重要影响,植物的繁殖能力与其扩散能力呈正相关 (Barret, 1983; 张斯斯和肖宜安, 2013)。许多外来植物在迁入地迅速繁殖,花期长、花数量多,能够吸引更多的授粉昆虫,而且不需要专化传粉者,产生更多种子,并且种子细小,或有的具有冠毛,容易传播(Reichard和Hamilton, 1997; 曾建军等, 2010)。有效的扩散机制和良好的集群能力,使这些植物能快速蔓延,成为入侵植物较普遍的类群之一。如一年蓬(Erigeronannuus)、钻形紫菀(Aster subulatus)等菊科植物(张斯斯和肖宜安, 2013)。风媒植物因无需生物传粉者,不存在传粉者缺乏而导致的自然授粉障碍,更容易实现入侵,如禾本科(Gramineae)植物具有有效的风媒传粉机制,许多种类能在新分布地迅速传播。虫媒植物在到达新分布区后往往因缺乏合适的传播媒介而影响其扩散能力 (Gasso等, 2009)。然而,大多数能够快速扩散的外来植物除了有性繁殖之外,克隆繁殖能力也非常强 (李瑞利等, 2007)。例如菊科剑叶金鸡菊(Coreopsis lanceolata)在多种生境下均能快速密集型克隆生长,其种群耐贫瘠和干旱能力非常强,因而适应不同生境的能力也非常强,这种生长策略使剑叶金鸡菊个体能快速扩增并排挤本土植物,形成大面积的单一优势种群 (曾建军等, 2012)。入侵植物豚草(Ambrosia artemisiifolia)也存在强烈的克隆繁殖现象 (Friedman和Barret, 2008)。外来植物在迁入地选择有性繁殖或无性繁殖策略,取决于迁入地是否存在有性繁殖的障碍,如传粉者缺乏、迁入地缺乏花器官发育所必需的光照、温度或其他条件等。张斯斯和肖宜安(2013)统计了入侵中国的137种植物的繁殖方式,发现其中克隆繁殖植物占了35.8%,但绝大多数为菊科和禾本科的草本植物。
在本研究中,对大花老鸭嘴的花特征、构造和传粉观测均显示该物种是典型的虫媒花物种。花序和居群花期较长,但单花花期短;花冠色彩艳丽,花冠两侧对称,具有花蜜(图1B、H),花气味清香,符合蜂媒花的一般特征 (Thomann等, 2013)。而且两侧对称的花由于能控制传粉者的访花姿态,优化了传粉者与花部结构之间的相互作用,大大提高了传粉效率 (张勃等, 2010)。扁柄木蜂是大花老鸭嘴最有效的传粉者,花冠切除实验结果显示:完全切除花冠的花,扁柄木蜂访花频率为0,而半切除和未切除花冠的花,访花频率最高时段均达到2-2.8 n·h-1(图5),说明大花老鸭嘴的淡紫色花冠对扁柄木蜂具有较强的吸引力。大花老鸦嘴花冠筒喇叭状,外宽内窄,柱头位于花冠喉部,向下弯曲,而扁柄木蜂体型与花冠筒内壁的大小接近,体表被密毛,在其入花筒取食花蜜时,前背与柱头相互触碰,从而使落在扁柄木蜂背上的花粉粘到柱头上,成功授粉。背部传粉使传粉者无法触及和梳理花粉而避免了浪费,从而保证了花粉的有效散播和高效利用(Ngamriabsakul, 2005)。

图6 花冠筒对扁柄木蜂访花频率的影响Fig. 6 Effect of corolla tubes for visiting frequency of Xylocopa latipes
另外,大花老鸭嘴在花冠筒基部分泌的花蜜(图1H、L),吸引传粉者的同时,也吸引了大量的蚂蚁,长期蛰伏于大花老鸭嘴的花序、花萼、花冠上(图1L)。这些蚂蚁能够清除蛾类、蝶类等在花冠或柱头上所产的卵及孵化的幼虫,防止幼虫对花朵的破坏 (Fiala等, 1996)。而且蚂蚁的群聚性、好斗性,使得其他昆虫不敢冒犯它们的领地(曹坤方, 1992),充当“护花使者”的角色。野外观测过程中发现在没有蚂蚁的花序上,花冠、雄蕊和雌蕊均会遭到毒蛾类幼虫的啃食,同时也有其他小昆虫栖息于花冠内部,由此可见,蚂蚁对大花老鸭嘴的花冠具有重要的保护作用。再者,大花老鸦嘴的4枚雄蕊略呈2长2短的排列,位于前面的长雄蕊的花药与位于后侧的短雄蕊的花药结构有一定的区别,长雄蕊的花药纵裂部位鬃毛明显浓密,而短雄蕊的鬃毛较为稀疏,在花药开裂和散粉过程中,这些鬃毛上布满花粉,传粉者扁柄木蜂背部也有浓密的毛被,在与花药接触过程中,头部或前胸触碰到长雄蕊,短雄蕊反向挤压长雄蕊,主要带走长雄蕊的花粉(图1J、K)。这种同一朵花内雄蕊和花药的结构和功能分化,也是虫媒花的重要特征之一(任明迅, 2009)。另外,大花老鸭嘴的柱头表面具有非常密集的短毛状突起(图2C),在扁柄木蜂携带花粉访花时,头部和背部的花粉会与大花老鸭嘴的柱头充分接触,短毛状突起的边缘可让足够多的花粉滞留在柱头上。光学显微镜下观测发现大花老鸭嘴的子房发育正常,不存在子房败育现象 (图1G)。这些研究结果均充分证明大花老鸭嘴具有虫媒花的典型特征。然而,连续3年在4个研究居群和以及其它踏查的居群内,均未发现大花老鸭嘴的自然结实,人工自花受粉、异株授粉等处理的结果率也均为0,说明大花老鸭嘴在研究区域内,不是通过种子生产来繁衍后代,而是通过无性繁殖,进行种群扩散。
3.2 大花老鸭嘴的繁殖策略推测
在中国华南地区的园林栽培植物中,也有不少只开花不结果的物种。比较常见的种类有白兰(Michelia alba)、大红花(Hibiscus rosa-sinensis)、悬铃花(Malvaviscus arboreus var. penduliflorus)和红花羊蹄甲(Bauhinia blakeana)等;栽培的苹果(Malus pumila)、樱桃(Cerasus pseudocerasus)等果树也存在只开花不结果的现象(原录山等, 2010; 艾提江, 2008)。导致植物只开花不结果的因素可分为内因和外因,其中内因包括多倍体、花粉败育、子房败育、花粉与柱头不亲和等;外因包括缺乏传粉昆虫、花期天气异常、土质过于肥沃或营养不足等。针对大花老鸭嘴只开花不结果的现象,本研究的结果显示其花粉活力正常 (图3),而且授粉后的花粉在柱头上能萌发出花粉管 (图2D),说明大花老鸭嘴的花粉和柱头发育正常,但花粉管在授粉后3 d尚未延伸至子房位置,说明花粉管在柱头和花柱上的生长发育可能存在障碍。而且大花老鸭嘴的花在凋谢后,雌蕊会自然脱落。如花粉管在3 d尚未延伸至子房,子房已随着雌蕊凋落,无法完成胚珠受精和结实。扁柄木蜂为大花老鸭嘴最有效的传粉者,访花频率和传粉效率也很高,说明大花老鸭嘴在迁入地不存在传粉者缺乏的有性繁殖障碍。另外大花老鸭嘴居群花期很长,全年均可开花,花期天气异常等外因的影响较小;而且大花老鸭嘴的生态适应性强,对生境土壤要求不严,沟边、房屋边、道路边均能健壮生长,但在这些生境均未发现其自然结果,故其只开花不结果的现象也不太可能是土质过于肥沃或营养不足所导致。根据以上研究结果,初步推测大花老鸦嘴可能存在花粉管在柱头和花柱上生长存在障碍,进而造成胚珠未能正常受精,造成所有植株不结果的现象。
大花老鸦嘴目前在华南地区大多数用于藤蔓绿化和垂直绿化,栽培方式主要为扦插和块根等无性繁殖,生长迅速 (黄芳, 2009),在夏威夷、新加坡、澳大利亚等地也是采取人工无性繁殖的方式,大面积繁殖(Daswon等, 2008)。通过扦插和块根繁殖的植株,其母株可能源自亲缘关系较近的亲本或无性系。这些亲本或无性系植株之间的花相互授粉,均可能产生“自交不亲和”现象,从而使得大花老鸭嘴每年长出大量的花,但却未见任何有性繁殖结出的果实。另外,染色体多倍化也可能造成大花老鸭嘴无法正常结实。有关大花老鸭嘴在迁入地采取借助人工无性繁殖的策略,随着人为活动快速扩散,而放弃可能更有利于后代适应新环境的有性繁殖方式,其最终原因尚需进一步广泛取样,对其染色体数目、核型开展研究。同时结合AFLP等分子标记方法,估算居群间基因渐渗现象、遗传多样性等,最终弄清其从有性走向无性的内在和外在原因。
4 结论
大花老鸭嘴的花具有典型的虫媒花特征,颜色鲜艳,两侧对称,花粉表面螺旋状萌发孔,方便传粉者将其在不同花之间传递,花粉、柱头和子房发育正常,最有效的传粉者为扁柄木蜂,传粉效率高,飞行距离远,可在居群内不同植株间和居群间相互传递花粉。说明大花老鸭嘴在迁入地不存在传粉者缺乏的障碍,而且花结构也满足异花授粉的条件,但研究区域内未发现其任何结果的现象。综合电镜扫描结果,初步推测华南地区的大花老鸭嘴植株可能因为长期的人工无性繁殖,其植株亲缘关系较近,遗传分化小,不同居群和植株之间相互授粉,可能仅仅是花粉和柱头之间“自交不亲和”,或“染色体多倍化”,这也许是只开花不结果的重要原因。
ABHILASHA D, JOSHI J. 2009. Enhanced fitness due to higher fecundity, increased defence against a specialist and tolerance towards a generalist herbivore in an invasive annual plant [J]. Journal of Plant Ecology, 2: 77-86.
BARRET S C H. 1983. Crop mimicry in weeds [J]. Economic Botany, 37: 255-282.
DAWSON W, BURSLEM DFRP, HULME P E. 2009. Factors explaining alien plant invasion success in a tropical ecosystem differ at each stage of invasion [J]. Journal of Ecology, 97: 657-665.
DAWSON W, MNDOLWA A S, BURSLEM DFRP, et al. 2008. Assessing the risks of plant invasions arising from collections in tropical botanical gardens [J]. Biodiversity and Conservation, 17: 1979-1995.
FIALA B, KREBS A, BARLOW H S, et al. 1996. Interactions between the climber Thunbergia grandiflora, its pollinator Xylocopa latipes and the ant Dolichoderus thoracicus: The "nectar-thief hypothesis" refuted? [J]. Malayan Nature, 50: 1-14.
FRIEDMAN J, BARRET S C H. 2008. High outcrossing in the annual colonizing species Ambrosia artemisiifolia (Asteraceae) [J]. Annals of Botany, 101: 1303-1309.
GASSO N, SOL D, PINO J, et al. 2009. Exploring species attributes and site characteristics to assess plant invasions in Spain [J]. Diversity and Distributions, 15: 50-58.
LEE C E. 2002. Evolutionary genetics of invasive species [J]. Trends in Ecology and Evolution, 17: 386-390.
MACK R N, SIMBERLOFF D, MARK L W, et al. 2000. Biotic invasions: causes, epidemiology, global consequences, and control [J]. Ecological Applications, 10: 689-710.
MEYER L. 2004. Beautes fatales: Acanthaceae species as invasive alien plants on tropical Indo-Pacific Islands [J]. Diversity and Distributions, 10: 333-347.
NGAMRIABSAKUL C. 2005. Morphological study of the versatile anther group in the tribe Zingibereae (Zingiberaceae) [J]. Walailak Journal of Science and Technology, 2: 11-12.
REICHARD S H, HAMILTON C W. 1997. Predicting invasions of woody plants introduced into North America [J]. Conservation Biology, 11: 193-203.
RICHARDSON D M, PYSEK P, REJMANEK M, et al. 2000. Naturalization and invasion of alien plants: concepts and definitions [J]. Diversity and Distributions, 6: 93-107.
THOMANN M, IMBERT E, DEVAUX C, et al. 2013. Flowering plants under global pollinator decline [J]. Trends in Plant Science, 18: 353-359.
艾提江. 2008. 庭院苹果长期不结果的原因及改良措施[J]. 新疆农垦科技, 5: 28-29.
曹坤方. 1992. 有花植物与传粉动物[J]. 植物杂志. 19: 27-30.
曾建军, 肖宜安, 孙敏. 2010. 入侵植物剑叶金鸡菊的繁殖特征及其与入侵性之间的关系[J]. 植物生态学报, 8: 966-972.
曾建军, 肖宜安, 周小军. 2012. 入侵植物剑叶金鸡菊克隆繁殖特性及群落特征[J]. 植物研究, 2: 147-150.
柴伟玲, 类延宝, 李扬苹, 等. 2014. 外来入侵植物飞机草和本地植物异叶泽兰对大气CO2浓度升高的响应[J]. 生态学报, 2014, 34(13): 3744-3751.
陈国菊, 刘厚成, 杨瑞陶, 等. 2001. 遮阴对大花山牵牛和桂叶老鸦嘴生长及叶片组织结构的影响[J]. 华南农业大学学报, 22: 56-59.
洪岚, 沈浩, 杨期和, 等, 2004. 外来入侵植物三叶鬼针草种子萌发与贮藏特性研究[J]. 武汉植物学研究, 5: 433-437.
黄芳. 2009. 观花藤本植物新秀——大花老鸦嘴[J]. 南方农业, 3: 20.
江纪武. 2003. 肯尼亚、印度等国民族药[J]. 国外医学:中医中药分册, 25: 119-123.
姜勇, 李艳红, 王文杰, 等. 2013. 光和不同打破种子休眠方法对紫茎泽兰种子萌发及幼苗状态的影响[J]. 生态学报, 1: 302-309.
李瑞利, 石福臣, 张秀玲, 等. 2007. 天津沿海滩涂互花米草种群生殖分株数量特征及生殖分配研究[J]. 植物研究, 27: 99-106.
陆耀东, 何松, 晓洪东, 等. 2008. 大花老鸦嘴组织培养技术[J]. 林业实用技术, 9: 43.
彭东辉, 张启翔, 黄启堂, 等. 2008. 多花野牡丹传粉生物学观察[J]. 福建林学院学报, 28: 115-120.
任明迅, 张全国, 张大勇. 2004. 入侵植物凤眼蓝繁育系统在中国境内的地理变异[J]. 植物生态学报, 28: 753-760.
任明迅. 2009. 花内雄蕊分化及其适应意义[J]. 植物生态学报, 33: 222-236.
吴冲, 史正军. 2008. 华南地区9种攀缘植物的季节性扦插繁殖研究[J].广西园艺, 19: 6-8.
杨东娟, 朱慧. 2008. 入侵植物南美蟛蜞菊克隆繁殖特性初探[J]. 安徽农业科学, 15: 6469-6470, 6532.
原录山, 刘万和, 张殿高. 2010. 大樱桃不结果的原因与对策[J]. 北方果树, 4: 32-33.
张勃, 孙杉, 张志强, 等. 2010. 杠杆状雄蕊及其进化生态学意义[J]. 植物生态学报, 34: 89-99.
张斯斯, 肖宜安. 2013. 中国外来入侵植物生活型与性系统多样性[J].植物研究, 3: 351-359.
钟军弟, 徐意媚, 曾富华, 等. 2014. 不同生境下假臭草生长特征分析[J]. 广西植物, 1: 68-73, 76.
Pollination and reproductive strategy of the alien plant Thunbergia grandiflora in the immigrated habitats
TANG Guangda*, CHENG Jianfei, XIE Zhenxing, CHEN Jing, LIN Cuiju
College of Forestry, South China Agricultural University, Guangzhou 510642, China
The survival and reproductive strategy of alien plants in new place have become the research hotspots recently. It is beneficial for exploring the survival and reproductive mechanisms of alien plants to investigate their breeding strategy, which will be useful for the reasonable exploitation of the alien plants. In this paper, the phenology, floral structure, pollen viability, pollinators and their behaviors of Thunbergia grandiflora were studied in four populations in Guangzhou and Shenzhen, Guangdong Province. The results showed that the florescence of single flower was one day only. The pollen viability of long stamen was the highest (73.92%) at 10: 00, and the short stamen is 71.54%, and it declined gradually after 12:00. The effective pollinator of T. grandiflora was Xylocopa latipes. The frequency of flower-visiting was higher (5 h-1) between 9: 00—10: 00 than any other flowering time, and the number of pollinator decreased after 13: 00 pm. No any effective pollinators visit flowers of T. grandiflora after 14:00 except for a few ants inhabiting on the florescence. Field experiments indicated that the fruit set of all six treatments, among artificially out-crossed treatment, self-pollinated treatment, and natural pollinated treatment, was 0%. The growth of ovary was normal under the observation of light microscope (LM). The result of scanning electronic microscope (SEM) showed that the pollens of T. grandiflora could germinate on the surface of stigma, but the pollen tubes were very short and could not grow into the ovary. The main artificial propagation way of T. grandiflora was the cloning with its tuberous roots and stems. Therefore, it was speculated that the plants of T. grandiflora coming from different populations in the study area might derive from the same or similar clones, and the genetic relationship of T. grandiflora are probably close between different populations. All out-crossed and self-pollinated flowers might be‘self-incompatiblility’ or ‘chromosomal polyploidization’. Therefore, it could not produce fruits in its immigrated habitats.
Thunbergia grandiflora; pollination biology; Xylocopa latipes; self-incompatibility
Q948
A
1674-5906(2014)06-0950-08
唐光大,成建飞,谢振兴,陈瑾,林翠菊. 外来植物大花老鸭嘴在迁入地的传粉机制和繁殖策略[J]. 生态环境学报, 2014, 23(6): 950-957.
TANG Guangda, CHENG Jianfei, XIE Zhenxing, CHEN Jin, LIN Cuiju. Pollination and reproductive strategy of the alien plant Thunbergia grandiflora in the immigrated habitats [J]. Ecology and Environmental Sciences, 2014, 23(6): 950-957.
广东省教育厅高校优秀青年创新人才培养计划项目(LYM11032)
唐光大(1982年生),男,讲师,博士,从事植物系统进化和传粉生物学研究,E-mail: gdtang@scau.edu.cn;*通信作者
2014-03-30
猜你喜欢
皖西学院学报(2022年5期)2022-10-26
河北农机(2021年11期)2021-11-29
野草(2020年4期)2020-08-13
江汉大学学报(自然科学版)(2019年6期)2019-12-06
孩子·小学版(2019年11期)2019-09-10
农业机械学报(2018年11期)2018-12-04
山西农业科学(2018年9期)2018-09-12
农业机械学报(2017年9期)2017-10-11
北方音乐(2017年5期)2017-04-13
生物学教学(2017年6期)2017-02-18
