
KCNJ11基因E23K基因多态对细胞膜电流的影响*
2014-01-22 10:00夏小慧杨爱宏
中国应用生理学杂志 2014年1期
夏小慧,杨爱宏,胡 扬
KCNJ11基因编码内向整流型钾离子通道蛋白Kir6.2,主要存在于心肌、骨骼肌、胰腺β-细胞、脑神经元等处[1,2]。细胞内 ATP/ADP的变化、4,5-二磷酸肌醇以及长链乙酰辅酶A均能激活该通道。在运动过程中,能合理调节心脏对外界应急反应的适应能力,被认为是心脏的能量感受器。
KCNJ11基因位于11p 15.1区域,仅1个外显子,无内含子,编码一个390个氨基酸的多肽。E23K多态(rs5219,C595T)是位于该基因外显子处的错义突变。该基因编码蛋白是一种ATP敏感性钾离子通道,与能量代谢密切相关。大量研究表明,E23K多态与糖尿病[3]、心血管疾病[4]密切相关。我们推测,E23K变异可能会改变蛋白质性质,从而改变钾离子通道的电流。在此假设前提下,本研究利用全细胞膜片钳技术,分析携带不同等位基因的重组质粒转染真核细胞后,对细胞膜表面电流密度的影响,以探讨E23K多态与相关疾病关联的机制。
1 材料与方法
1.1 材料
质粒提取试剂盒、PCR试剂盒购自sangon公司;Taq聚合酶、Pfu酶、EcoR I、Xba I限制性内切酶等购自Progema公司;Lipofectamine 2000购自Invitrogen公司;DMEM细胞培养液、胎牛血清、青霉素、胰蛋白酶等购自 Giboc公司;氯化钠、氯化钾、EDTA、HEPES、氯化镁、D-葡萄糖等购自Sigma公司;其他试剂均为国产分析纯。
感受态细胞TOP10购自Novasygen公司,pcDNA3.1/CT-GFP质粒载体由兰州兽医研究所人兽共患病研究室惠赠。健康人基因组DNA由本课题组前期保存。
1.2 KCNJ11基因的扩增及表达载体的构建
根据已发表的 GenBank(NCBI Reference Sequence:NM-000525.3)中的 KCNJ11序列设计引物F、R,上下游引物 5'端分别加载 EcoR I和 Xba I酶切位点和保护碱基。以健康人基因组DNA为模板,PCR扩增KCNJ11基因外显子。将扩增产物纯化后,通过连接、转化后插入pcDNA3.1/CT-GFP载体。测序得到多态位点为G(对应于氨基酸E)的重组载体。载体命名为 pcDNA3.1-KCNJ11(E)(图 1)。

Fig.1 Restriction endonuclease digestion confirmations of recombinant plasmid1:Digestion fragments by EcoR I and Xba I;M:DNA marker
根据突变位点,设计突变引物Fm和Rm。以pcDNA3.1-KCNJ11(E)为模板,以 F和 Fm、R和 Rm引物对分别进行PCR扩增,所得PCR产物I和产物II经纯化后等比例混合。取混合物为模板,以F和R引物对进行扩增,纯化回收PCR产物,经连接、转化后插入 pcDNA3.1/CT-GFP载体。测序得到多态位点为A(对应于氨基酸K)的重组载体。载体命名为pcDNA3.1-KCNJ11(K)。本研究所用引物序列见表1。

Tab.1Primers for amplification
1.3 细胞转染
HEK293T细胞在 DMEM培养基中(含 10%FBS),37℃,5%CO2,100%相对湿度条件下常规培养。当细胞浓度达2×105cells/ml时,用重组质粒(除去内毒素)pcDNA3.1-KCNJ11(K)、pcDNA3.1-KCNJ11(E)分别转染细胞。转染试剂为 Lipofectamine 2000,转染过程严格按照转染说明书进行。转染后24~48 h在荧光显微镜下观察细胞转染效率,选边缘整齐、表面光滑,有绿色荧光的细胞进行电生理测定。
1.4 电生理记录
配制细胞外液(mmol/L:NaCl:140,KCl:5.4,Ca:1.8,MgCl2:0.5,NaH2PO4:0.33,HEPES:5,葡萄糖:5.5,用 NaOH调 pH 7.4)、细胞内液(K:130,MgCl2:1,磷酸肌酸二钠盐:5,天冬氨酸:90,EGTA:10,HEPES:5,用 KOH调 pH 7.4)。通过三维操纵仪(MP285R,Sutter)使电极与细胞表面形成高阻封接及全细胞记录模式,电流信号经Axon 700B放大器(Axon)进行处理。钳制电压为由-150 mV至60 mV,阶跃电压:10 mV,采样时间:250 ms。采样频率:5 KHz。用pClamp9软件(Axon)进行数据分析。由电流振幅与电容的比值计算电流密度(pA/pF)。数据以平均数±标准差表示。对两组数据进行T检验。
2 结果
2.1 目的基因的获得和表达载体的构建
成功扩增到大小为1 173 bp的片段,并将其插入pcDNA3.1载体。测序结果表明,与GenBank公布的KCNJ11基因序列相比,同源性为99.6%(图1)。
图 2所示为构建的 pcDNA3.1-KCNJ11(E)、pcDNA3.1-KCNJ11(K)重组质粒部分测序图,箭头所示为突变位点。结果表明,构建的两个载体仅有突变位点差异,其余序列均一致。
2.2 细胞转染
细胞转染结果表明,重组质粒成功转染入HEK293T细胞,并在细胞内表达,平均转染率为13.6%。内有绿色荧光者表示已转染并KCNJ11基因成功表达的细胞(图3)。

Fig.2 Sequenced recombinant plasmid fragments
Fig.3 HEK293T cells after transfected
2.3 细胞膜电流密度测定
全细胞膜片钳记录结果显示,pcDNA3.1-KCNJ11(E)转染后的细胞中记录到了相对较强的电流,而pcDNA3.1-KCNJ11(K)转染后的细胞中记录到的电流明显弱(图4)。
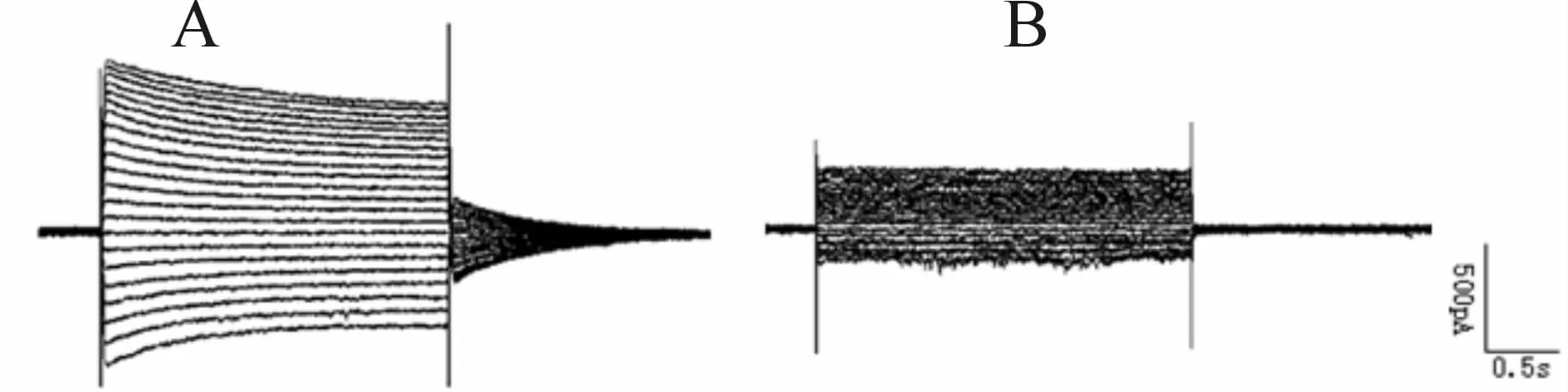
Fig.4 Currents in transfected cell was investigated using patch clampA:pcDNA3.1-KCNJ11(E);B:pcDNA3.1-KCNJ11(K)
当钳制电压为由-150 mV渐变为+60 mV,根据电流随电压的变化绘制电压电流关系曲线(图5)。电流正值为外向电流,电流负值为内向电流。在两种不同质粒转染后的HEK 293T细胞中,其细胞膜的反转电位约为-50 mV,内向电流与外向电流大小相近。而在pcDNA3.1-KCNJ11(K)转染后的细胞中,内向电流和外向电流均明显减少。T-检验结果显示,两组细胞表面电流对比有显著性差异(P<0.05)。
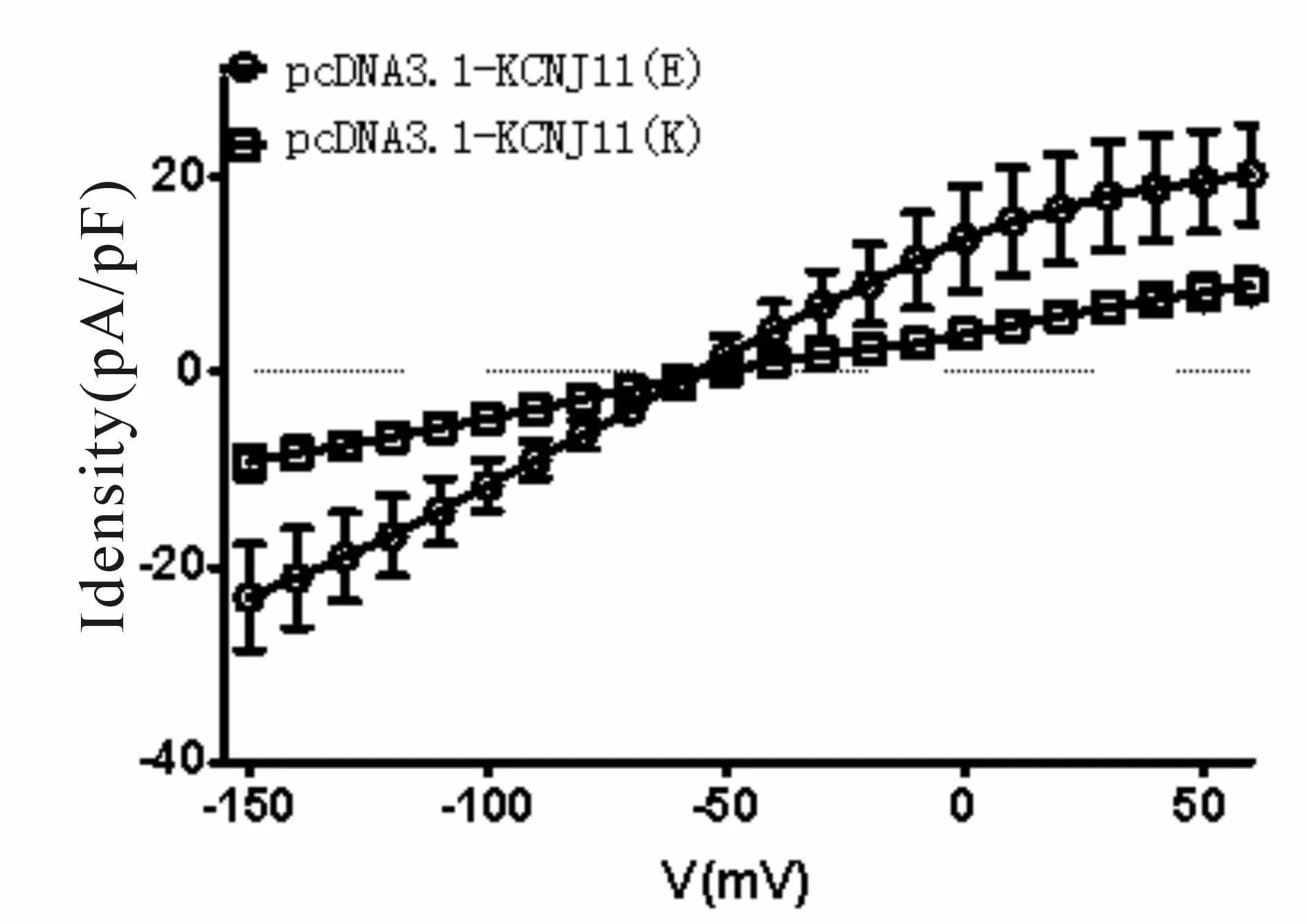
Fig.5 I/V curves of ATP-sensitive K+(KATP)channels after transfected difference recombinant plasmids(n=10)
3 讨论
KCNJ11基因编码的Kir6.2蛋白分布于心肌、骨骼肌、平滑肌、内分泌细胞等,其生理功能是维持细胞静息电位,调节骨骼肌、血管平滑肌的舒缩等,并参与多种生理功能:如胰腺胰岛素的分泌、心肌缺氧的保护功能等。钾离子通道的特征是其开放活性受胞内ATP的调节,ATP及磺酰脲类药物抑制其开放,而ADP及其它二磷酸核苷促其开放。KCNJ11基因编码蛋白的通道上,发现许多影响ATP敏感性的氨基酸位点和区域[5],如 R50、K185、R201、G334等四个位点共同形成一个ATP结合槽,其中任何一个氨基酸的改变会使通道对ATP的敏感性极大降低,甚至消失。
体外表达系统是研究离子通道电流特征的一个重要途径[6],而膜片钳技术被认为是研究离子通道的“金标准”。离子通道的特征,一定程度上依赖于使用的表达系统。如植物钾离子通道蛋白KAT1在蛙卵中的激活电位接近-80 mV,而在昆虫细胞中的接近-60 mV[7]。本实验中,测到的细胞膜电流较小,最大为20 pA/pF,最小为-23 pA/pF。这可能与表达载体的类型、转染细胞以及实验条件有关。由于本研究两组转染质粒只有一个位点差异,其余条件均一致,因此对比两组转染质粒的电生理功能,结果可靠。
E/K突变是KCNJ11基因中研究最多的多态位点。大多数研究认为该位点多态与糖尿病相关联,但结果又不尽一致[8,9]。对各独立研究进行合并统计分析,提示KCNJ11基因KK基因型可能是Ⅱ型糖尿病发病的危险因子[10]。通过构建分子模型显示,Kir6.2通道结构中,E23残基在胞内功能性的α-螺旋区,而E残基被K取代后,该氨基酸残基带有相反的电荷,有可能导致结构重排以及Kir6.2亚基相邻的内部结构断裂,特别是在纯合子状态下,这种作用更为明显。也有研究认为,由于KATP通道是由一个内向整流性钾通道(Kir6.2或Kir6.1)和一种 ATP结合蛋白—磺酰脲类受体(SUR1或SUR2)组成的异聚体[11],在 ABCC8基因(编码 SUR1蛋白)上有一个敏感的S1369A多态位点,是KCNJ11基因E23K位点与ABCC8基因S1369A位点共同作用改变了离子通道的ATP敏感性[12]。还有研究认为,钾离子通道电流的变化依赖于ABCC8中A1369变异的存在,而不是KCNJ11中K23[7]。而在本研究中,两个转染质粒只有E/K位点不同,其余条件均相同,而检测结果细胞表面电流发生了明显变化,说明E23K多态位点单独可以改变离子通道的功能,从而改变细胞相关特性。
总之,人 KCNJ11基因 E23K通过异源表达载体,转染293T细胞后,细胞表面电流发生了明显改变,表明KCNJ11基因外显子E23K多态能导致细胞膜电流发生改变。为进一步研究多态位点与相关疾病的关联机制提供实验基础。
[1] Banas K,Clow C,Jasmin B J,et al.The KATPchannel Kir6.2 subunit content is higher in glycolytic than oxidative skeletal muscle fibers[J].Am J Physiol Regul Integr Comp Physiol,2011,301(4):R916-925.
[2] 陈 悦,徐 晓,鲁 颖.葛根素对大鼠心室肌细胞动作电位及钾通道电流的影响[J].中国应用生理学杂志,2010,26(2):246-248.
[3] Li YY.The KCNJ11 E23K gene polymorphism and type2 diabetes mellitus in the Chinese Han population:a meta-analysis of 6,109 subjects[J].M Biol Rep,2013,40(1):141-146.
[4] Yi Y,Dongmei L,Phares D A,et al.Association between KCNJ11 E23K genotype and cardiovascular and glucose metabolism phenotypes in older men and women[J].Exp Physiol,2008,93(1):95-103.
[5] Trapp S,Proks P,Tucker SJ,et al.Molecular analysis of ATP-sensitive K channel gating and implications for channel inhibition by ATP[J].J Gen Physiol,1998,112(3):333-349.
[6] 罗王倩,崔文玉,汪 海.SH-EP1细胞株转染表达神经元 N受体α4β2、α4β4和α7亚型激活态的电生理学特征[J].中国应用生理学杂志,2009,25(3):392-396.
[7] Marten I,Gaymard F,Lemaillet G,et al.Functional expression of the plant K+channel KAT1 in insect cells[J].FEBSLett,1996,380(3):229-232.
[8] Alsmadi O,Al-Rubeaan K,Wakil SM,et al.Genetic study of Saudi diabetes(GSSD):significant association of the KCNJ11 E23K polymorphism with type 2 diabetes[J].Diabetes Metab Res Rev,2008,24(2):137-140.
[9] Babenko A P,Aguilar-Bryan L,Bryan J.A view of sur/KIR6.X,KATP channels[J].Annu Rev Physiol,1998,60:667-687.
[10] Hamming K S,Soliman D,Matemisz L C,et al.Coexpression of the type 2 diabetes susceptibility gene variants KCNJ11 E23K and ABCC8 S1369A alter the ATP and sulfonylurea sensitivities of the ATP-sensitive K+channel[J].Diabetes,2009,58(10):2419-2424.
[11]田 芸,李 亚.β-细胞腺苷三磷酸-敏感性钾通道基因E23k多态性与2型糖尿病关系的Meta分析[J].中国全医科学,2010,13(20):2233-2235.
[12] Schwanstecher C,Meyer U,Schwanstecher M.K(IR)6.2 polymorphism predisposes to type 2 diabetes by inducing overactivity of pancreatic beta-cell ATP-sensitive K+channels[J].Diabetes,2002,51(3):875-879.
猜你喜欢
世界科学技术-中医药现代化(2022年2期)2022-05-25
世界科学技术-中医药现代化(2021年12期)2021-04-19
心肺血管病杂志(2020年5期)2021-01-14
新世纪智能(高一语文)(2020年12期)2020-06-01
听力学及言语疾病杂志(2019年1期)2019-01-14
电子技术与软件工程(2017年8期)2017-05-10
科技视界(2016年27期)2017-03-14
中国医疗美容(2015年4期)2015-04-27
科学中国人(2015年13期)2015-02-28
食品工业科技(2014年23期)2014-03-11
