
巍山红雪梨PpPAL基因的克隆及生物信息学分析
2014-01-19 06:54辛培尧唐军荣原晓龙孟富宣
经济林研究 2014年1期
董 娇,周 军,辛培尧,唐军荣,陶 磅,原晓龙,孟富宣
(1.西南林业大学 西南山地森林保育与利用省部共建教育部重点实验室,云南 昆明650224;2.中华全国供销合作总社昆明食用菌研究所,云南 昆明 650223;3.云南省农业科学研究院 园艺作物研究所,云南 昆明 650205)
苯丙氨酸解氨酶(phenylalanine ammonialyase,PAL)广泛存在于植物中,催化L-丙氨酸脱氨生成反式肉桂酸,在苯丙氨酸代谢途径中,它既是一种限速酶,也是一种诱导酶[1]。PAL在初生代谢和苯丙烷类代谢途径中起纽带作用[2]。PAL由4个亚基组成,由多基因家族编码,在这个多基因家族中,不同成员的表达特异性不同。不同植物中PAL基因的核苷酸序列与其编码的氨基酸序列的相似性较高,该基因家族可能起源于一个古老的基因,基因的复制和分子上的歧异可能会形成功能不同的基因,以应对不同环境因素的影响,分别在不同组织和发育时期表达[3]。有关研究结果表明,PAL与类黄酮、木质素和香豆素等次生代谢途径密切相关[4-7]。Koukol等人[8]首次从植物中发现并分离纯化了该酶。随着科学技术的发展及分离纯化技术的不断完善,对PAL的研究已迅速展开,目前已成功地从水稻、小麦、马铃薯、松树、豌豆等种植物中分离纯化得到PAL基因[9]。
巍山红雪梨是云南特有的红皮梨,笔者利用特异性引物对巍山红雪梨的PAL基因片段进行了同源克隆,并利用生物软件对该基因及其相应蛋白质进行了生物信息学分析,以期为调控巍山红雪梨类黄酮的积累及增强巍山红雪梨对不良环境的抵御能力的深入研究而奠定基础。
1 材料与方法
1.1 试验材料
1.1.1 植物材料
巍山红雪梨栽培于云南省安宁市清水河红皮梨种质基地,于2011年3月采集幼叶,经液氮处理后,保存于-80℃的冰箱中以备用。
1.1.2 主要试剂
反转录试剂盒(Quantscript RT Kit)、DNA割胶回收试剂盒、2× Taq PCR MasterMix Kit与DL marker 2000,均购自天根生物技术有限公司;常规化学药品及试剂,购自Sigma公司或国产AR级试剂;PCR和测序引物,由上海生工生物工程技术服务有限公司合成;测序由北京中美泰和有限公司完成。
1.2 试验方法
1.2.1 RNA的抽提与cDNA第一链的合成
RNA的提取:①取供试材料0.5~1.0g置于液氮中研磨成粉末,并快速将其转移到事先预冷的离心管中;②加入65℃预热的CTAB缓冲液 750μL 和β-巯基乙醇 60μL,于 65℃下处理5min,冷至室温再加入乙酸钾500μL,摇匀,于4℃下以12 000r/min的转速离心15min;③取上清液置于新管中,加入300μL的饱和酚,再加入24∶1的氯仿/异戊醇300μL,摇匀,于4℃下以12 000r/min的转速离心15min;④取上清液置于新管中,加入24∶1的氯仿/异戊醇600μL,摇匀,于4℃下以12 000r/min的转速离心15min;⑤取上清液置于新管中,加入体积分数为1/4的LiCl 4 mol/L,混匀,在4℃下沉淀过夜;⑥于4℃下以12 000r/min的转速离心20min,去上清液,用500μL的SSTE溶解沉淀后,加入水饱和酚250μL和24∶1的氯仿/异戊醇250μL,摇匀,于4℃下以12 000r/min的转速离心5min;⑦取上清液置于新管中,加入24∶1的氯仿/异戊醇500μL,摇匀,于4℃下以12 000r/min的转速离心5min;⑧取上清液置于另一新管中,加入无水乙醇1 000μL,于-80℃下沉淀30min;⑨于4℃下以12 000r/min的转速离心10min,弃上清液,用75%乙醇清洗沉淀物1次,在超净工作台上无菌风干燥至少15min,然后加入经灭菌处理过的DEPC水30μL溶解沉淀物,于-80℃的冰箱中保存以备用。
总RNA质量检测:取2μL的总RNA,用1%的琼脂糖凝胶电泳进行检测。
紫外分光光度计定量测定和纯度分析:取2μL RNA 样品稀释50倍,用紫外分光光度计测定230、260和280nm波长下的OD值,记录RNA浓度和OD260nm/OD280nm、OD260nm/OD230nm的值,以确定提取的巍山红雪梨的纯度。
cDNA第一条链的合成:①在冰浴的无核酸酶的离心管中依次加入1~5μg总RNA、2μL Oligo(dT)15、2μL dNTP(2.5 mM each)等 反应混合物,补RNase-free ddH2O定容至14.5μL;②于70℃下加热5min后迅速在冰上冷却2min,简短离心收集反应液后加入5×First-Strand Buffer(含有DTT)4μL和RNasin 0.5μL;③加入1μL(200 U)TIANScript M-MLV,轻轻地用移液器混匀;④于42℃下温浴50min;④于95℃下加热5min,终止反应,置之于冰上进行后续实验,或于-20℃下冷冻以保存;⑥用RNase-free ddH2O将反应体系稀释到50μL,取2~5μL进行PCR扩增反应。
1.2.2 巍山红雪梨PAL基因的克隆
引物设计:对GenBank上已经登录的梨PAL同源基因的多个序列进行比对,设计了3对特异性引物,引物序列分别如下。
PAL(F1):5’-CAGAACGGTGCTGTGGAGTC-3’;
PAL(R1):5’-GTGCTTCAACTTGTGCGTCA-3’;
PAL(F2):5’-TGGCATCTTATTGTTCCG-3’;
PAL(R2):5’-TTGGCTCACCGTGTTCTT-3’;
PAL(F3):5’-CACGGTGAGCCAAGTCG-3’;
PAL(R3):5’-GCATTCCGCAATCCTGT-3’。
PCR反应体系:以反转录得到的cDNA作为模板进行PCR扩增,其反应体系为cDNA模板2μL、2×PCR Master Mix 25μL、PAL(F)2μL、PAL(R)2μL、以 ddH2O 补足 50μL。
PCR反应程序:94℃,预变性2min;94℃、30s,56℃、30s,72℃、1min;35 个循环;72℃,延伸 7min。
反应结束后,以1%琼脂糖凝胶电泳检测PCR产物,将样品送至北京中美泰和有限公司进行测序。
1.2.3 巍山红雪梨PAL基因序列的生物信息学分析
测序由北京中美泰和有限公司完成。利用http://www.ncbi.nlm.nih.gov/、http://www.cbs.dtu.dk/、http://www.expasy.org/等网站提供的各类生物信息学软件进行在线分析。在NCBI-ORF Finder上进行开放阅读框(Open reading frame,ORF)的查找和翻译;利用ProtParam在线工具分析CHS的核酸及氨基酸序列的组成成分及其理化性质;利用MEGA4.0[10]完成氨基酸序列的同源性比对及进化树的构建;分别利用TargetP1.1(http://www.cbs.dtu.dk/services/TargetP/)、SignalP3.0(http://www.cbs.dtu.dk/services/SignalP/)、TMHMM2.0(http://www.cbs.dtu.dk/services/TMHMM/) 和ProtScale(http://www.expasy.ch/tools/ protscale.html)等在线工具进行蛋白质导肽与信号肽的预测、跨膜结构域及亲水性/疏水性的预测;分别利 用SOPMA(http://npsa-pbil.ibcp.fr/cgi-bin/npsa_automat.pl?page=/NPSA/npsa_sopma.html)、Swiss-Model(http://www.expasy.ch/swissmod/SWISSMODEL.html)在线工具和NCBI提供的Conserved Domain Search(CD-Search)服务器(http://www.ncbi.nlm.nih.gov/Structure/cdd/wrpsb.cgi)进行蛋白质二级、三级结构的预测及蛋白质功能域的分析。
2 结果与分析
2.1 PAL基因序列及其所推导的氨基酸序列的比对分析
经PCR扩增后,获得了3个片段,测序后将此3个序列进行拼接,获得了如图1所示的cDNA片段,该片段长为1 919 bp,编码628个氨基酸。将此序列进行比对分析,结果表明:其与西洋梨的PAL基因(DQ230992)、鸭梨Pyrus bretschneideri的PAL基因(GU906268、JQ247318)在核苷酸水平上的同源性分别为98%和95%、95%;其与甜樱桃Prunus avium的PAL基因(AF036948)、覆盆子Rubus idaeus的PAL基因(AF237955)、桑树Morus alba的PAL基因(HM064433)、荔枝Litchi chinensis的PAL基因(FJ944018)、葡萄的PAL基因(EF192469)和木薯Manihot esculenta的PAL基因(AY036011)的同源性分别为88%、83%、82%、80%、79%与78%。这一比对结果可以初步说明,PAL是苯丙氨酸解氨酶基因,名之为PpPAL,将此序列提交至GenBank,序列号为JQ749640。
根据巍山红雪梨PAL基因编码的氨基酸序列与已知的PAL基因的氨基酸序列,用Clustal W软件进行比对后,再用MEGA4.0软件采用NJ法作出它们的系统进化树,结果如图2A与B所示。从图2A中可以看出,巍山红雪梨的PAL蛋白与鸭梨(ADF59061、AEZ01784)、西洋梨(ABB70117)、甜樱桃(AAC78457)、覆盆子(AAF40224)、桑树(ADI40166)、荔枝(ACR15762)、葡萄(ABM67591)、木薯(AAK62030)、拟南芥(NP_001190223)的PAL蛋白聚在双子叶植物这一大类群中。在这个大类群中,同属植物巍山红雪梨、西洋梨和鸭梨都聚在一个亚类上,这说明它们的亲缘关系较近,这与从分类学上分析的结论相符;拟南芥的PAL蛋白和巍山红雪梨的PAL蛋白虽然聚在这个大类群中,但拟南芥在这个类群的外侧,说明二者在进化上的亲缘关系较远;单子叶植物玉米(NP_001241797)和水稻(AAO72666)聚为一类,说明它们的亲缘关系较近,但巍山红雪梨的PAL蛋白与它们之间在进化上的关系较远。分析序列比对结果后可以发现,此PAL序列包含了L180、V181、L230、A231等脱氨基氨基酸残基,其催化活性位点分别为N234、G235、N356、D357、N358、H370、HNQDV(460~464),这些活性位点与有关研究报道的水稻、玉米、夹竹桃、银杏的相同,说明PpPAL是PAL蛋白质家族成员之一。
2.2 PAL蛋白的理化性质
采用ProtParam在线软件分析PAL蛋白的理化性质,结果显示:PAL基因编码的蛋白质的预测分子量为68 KDa,理论等电点为6.42,其中Leu、Ala、Ser、Val等是其主要的氨基酸;负电荷残基(Asp+Glu)总数为66个,正电荷残基(Arg+Lys)总数为61个,不稳定系数为33.58,单条肽链属于稳定蛋白。

图1 巍山红雪梨PAL基因的核苷酸序列及由其推导的氨基酸序列Fig.1 Full-length sequences of PpPAL cDNA in Pyrus pyrifolia and its deduced amino acid sequence

A 巍山红雪梨的PAL蛋白与其它植物的PAL蛋白的进化关系A Evolutionary relationship of deduced PpPAL protein with PAL proteins from other plants
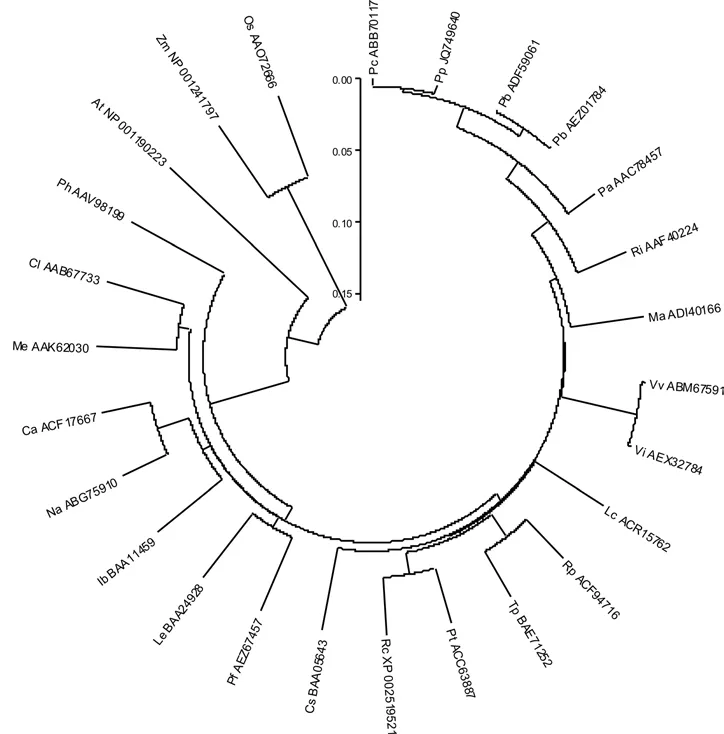
B 不同植物PAL蛋白的进化关系B Evolutionary relationship of PAL proteins in different plants
2.3 巍山红雪梨PAL蛋白的亚细胞定位、信号肽、跨膜结构和疏水性分析
采用Target 1.1 Server在线软件对PAL蛋白质亚细胞定位进行预测,结果显示:该蛋白质可能定位于细胞质基质,预测的可靠性等级为2级(详见表1),而与之同源性较高的其它植物(包括西洋梨、鸭梨等)的PAL基因也定位在细胞质基质上,相同的亚细胞定位说明这些基因可能具有相似的功能。

表1 巍山红雪梨PAL蛋白的亚细胞定位Table 1 PAL protein subcellular localization prediction of ‘Weishan’red skin pear (Pyrus pyrifolia Naki)
利用SignalP4.0 Server软件进行在线分析,结果也表明,巍山红雪梨的PAL基因编码的蛋白质无信号肽。可见,巍山红雪梨的PAL蛋白在游离核糖体上起始合成后,很可能没有进行蛋白质转运,而是继续保留在细胞质基质中催化L-苯丙氨酸脱氨生成反式肉桂酸。
利用TMHMM 2.0 Server软件对巍山红雪梨PAL氨基酸序列的跨膜结构域进行在线预测,结果表明,PAL整条肽链都位于细胞膜外,说明巍山红雪梨的PAL基因编码的多肽不存在跨膜区,结合上述转运肽的预测结果可以推测出,巍山红雪梨的PAL在细胞质基质中合成后很可能不经蛋白转运便直接锚定于细胞质基质中的特定部位而行使催化功能。
利用ProtScale软件对巍山红雪梨的PAL氨基酸序列的疏水性/亲水性进行预测,结果如图3所示。图3的预测结果表明,多肽链第322位具有最低分值-2.656,说明其亲水性最强;第256位具有最高分值(2.700),说明其疏水性最强。而就其整体而言,亲水性氨基酸均匀分布在整个肽链中,而且多于疏水性氨基酸。因此,整个多肽链表现为亲水性,没有明显的疏水区域。结合跨膜结构域的预测结果可以推断,巍山红雪梨的PAL基因编码的多肽不存在明显的疏水区域,这与PAL不存在跨膜区的特征相吻合。因此认为,巍山红雪梨的PAL是亲水性蛋白。

图3 巍山红雪梨PAL的疏水性/亲水性的预测结果Fig.3 Prediction of hydrophobicity or hydrophilicity of PAL in Pyrus pyrifolia
2.4 巍山红雪梨PAL蛋白的二级和三级结构的预测分析
利用SOPMA软件对巍山红雪梨的PAL氨基酸序列的二级结构进行预测,结果如图4(见封二)所示。图4表明,巍山红雪梨的PAL基因所编码的蛋白由57.39%的α-螺旋(Alpha helix)、7.79%的β-折叠(Extended strand)、4.61%的β-转角(Beta turn)和30.21%的随机卷曲(Random coil)组成。α-螺旋和无规则卷曲是巍山红雪梨PAL蛋白质二级结构的主要结构元件,β-转角和延伸链散布于整个蛋白序列中。
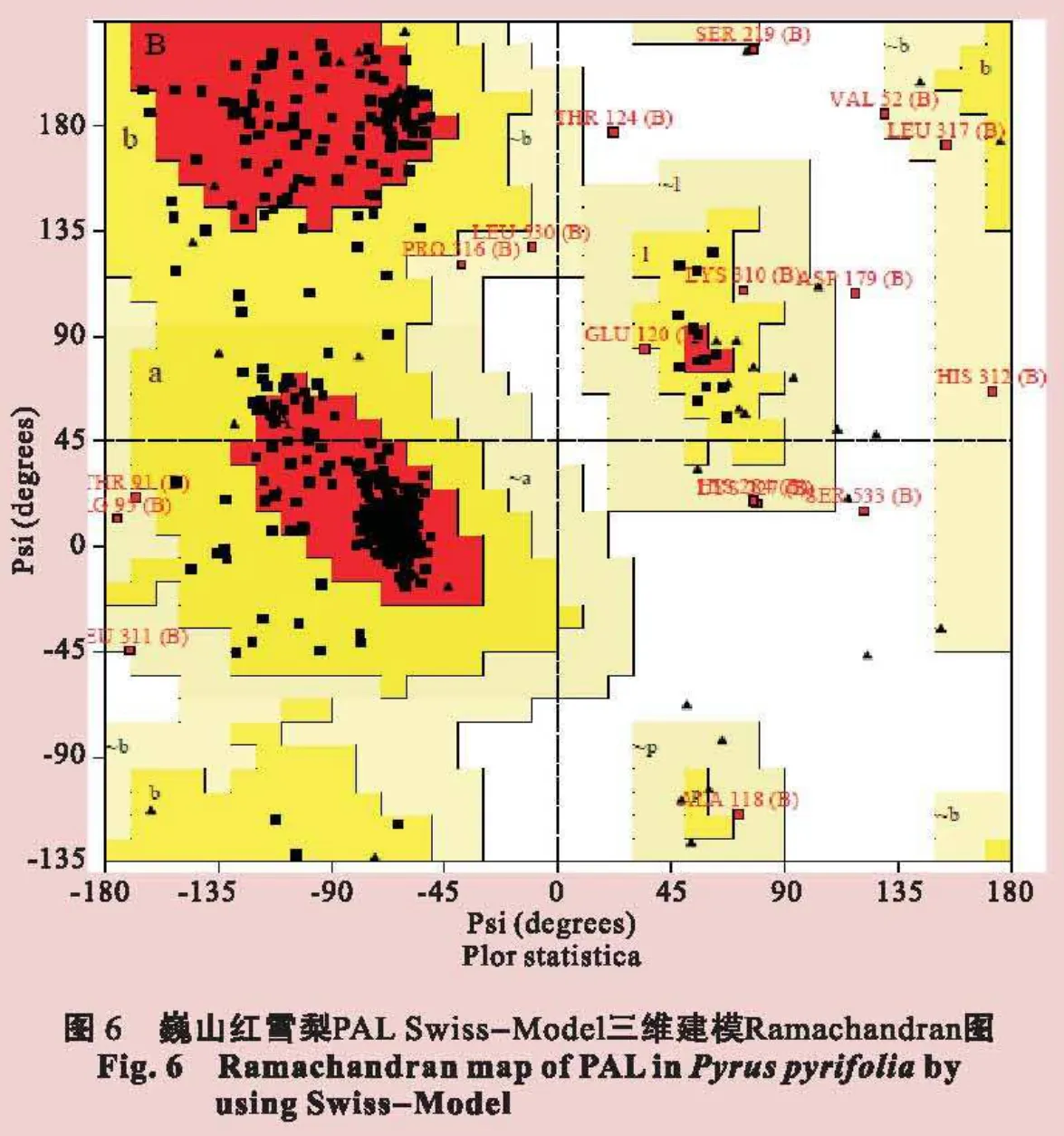
采用SWISS-MODEL同源建模的方法预测巍山红雪梨PAL蛋白的三级结构,结果如图5(见封二)所示。再利用ProCheck软件对建模结果进行检测,计算得出了如图6(见封二)所示的Ramachandran 图。图6表明,检测到的巍山红雪梨PAL蛋白质残基的二面角只有87.7%位于黄色区域,表明其空间结构可能不够稳定。
2.5 巍山红雪梨PAL蛋白结构功能域分析
使用NCBI提供的Conserved Domain Search service(CD-Search)对巍山红雪梨PAL蛋白功能域进行预测,结果如图7(见封二)所示。图7表明,该蛋白含有苯丙氨酸解氨酶-组氨酸解氨酶(PAL-HAL)和苯丙氨酸解氨酶(phe_am_lyase),它们都属于类裂解酶超家族(Lyase_I_like superfamily),在β-消除反应中起催化作用。
3 讨 论
苯丙氨酸解氨酶(PAL)是催化苯丙烷类代谢中的第一个酶。Neish早在1960年就证实了PAL催化花色素苷的合成[11]。此外,它在类黄酮、木质素、香豆素、水杨酸、单宁等次生代谢产物合成过程中也起着很重要的作用。这些次生物质对植物生长发育、抗病害、防御紫外线等方面都有帮助。近年来,有关PAL基因的研究也成为了相关研究的热点。
本研究从云南特有红皮梨品种——巍山红雪梨中克隆了PAL基因的cDNA片段,核苷酸与氨基酸序列分析结果表明,该基因与其它植物的PAL基因有着很高的同源性,并且含有相似的蛋白质保守活性位点。在PpPAL蛋白序列中包含L180、V181、L230、A231等脱氨基氨基酸残基,其催化活性位点分别为N234、G235、N356、D357、N358、H370、HNQDV(460-464),这些活性位点与对水稻[12]、玉米[13]、夹竹桃[14]等研究报道的结果相同。对PAL的聚类分析结果显示,巍山红雪梨与西洋梨、鸭梨的关系最近,处于同一分支,而与十字花科的拟南芥的进化关系稍远,与单子叶植物玉米、水稻的亲缘关系最远。本试验虽然只得到了PAL基因的片段,但克隆的基因具有代表性,为进一步克隆巍山红雪梨PAL基因全长奠定了基础[15]。
开展植物花色素苷合成调控机制的研究,除了利用模式植物,在一些如巍山红雪梨的木本植物中同样有利于揭示植物次生代谢途径在类黄酮积累中的调控规律。本研究结果可为红皮梨类黄酮类化合物代谢的分子调控机制及增强巍山红雪梨对不良环境的抵御能力的深入研究提供一定的理论依据。



[1] 谭洪花,曹尚银,房经贵,等.枣果实中苯丙氨酸解氨酶基因的克隆及序列分析[J].经济林研究,2011,29(1):15-20.
[2] 程水源,杜何为,许 锋,等.银杏苯丙氨酸解氨酶基因的克隆和序列分析[J].林业科学研究,2005,18(5):573-577.
[3] 凌文华,郭红辉.植物花色苷[M].北京:科学出版社,2009.
[4] DIXON R A,PAIVA N L.Stress-induced phenylpropanoid metabolism [J].The Plant Cell,1995,7:1085-1097.
[5] COCHRANE F C,DAVIN L B,LEWIS N G.TheArabidopsisphenylalanine ammonia lyase gene family: kinetic characterization of the four PAL isoforms [J].Phytochemistry,2004,65(11):1557-1564.
[6] FERRER J L,AUSTIN M B,STEWART C.Structure and function of enzymes involved in the biosynthesis of phenylpropanoids [J].Plant Physiology and Biochemistry,2008,46 (3):356-370.
[7] 江汉民,王 楠,赵 换,等.花椰菜苯丙氨酸解氨酶基因的克隆及黑腐病菌胁迫下的表达分析[J].南开大学学报:自然科学版,2012,45(4):87-92,98.
[8] KOUKOL J,CONN E E.The metabolism of aromatic compounds in higher plants IV Purification and properties of the phenylalanine deaminase ofHordeum vulgare[J].Journal of Biological Chemistry,1961,236:2692-2698.
[9] NI Y,JIANG H L,LEI B.Molecular cloning characterization and expression of two rapeseed (Brassica napusL)cDNAs orthologous toArabidopsis thalianaphenylalanine ammonia lyase [J].Euphytica,2008,159:1-16.
[10] TAMURA K,DUDLEY J,NEI M,et al.MEGA4: Molecular Evolutionary Genetics Analysis (MEGA)software version 4.0[J].Molecular Biology and Evolution,2007,24:1596-1599.
[11] NEISH A C.Biosynthesis pathway of aromatic compounds [J].Annual Review of Plant Physiology,1960,(11):15.
[12] MINAMI E,OZEKI Y,MATSUOKA M.Structure and some characterization of the gene for phenylalanine ammonia-lyase from rice plants [J].Eur J Biochem,1989,185(1):19-25.
[13] ROSLER J,KREKEL F,AMRHEIN N.Maize phenylalanine ammonia-lyase has tyrosine ammonia-lyase activity [J].Plant Physiol,1997,113(1):175-179.
[14] 王 燕,许 锋,杜何为,等.夹竹桃苯丙氨酸解氨酶的基因克隆与序列分析[J].华北农学报,2007,22(4):19-24.
[15] 孙百灵,王 旭,曲柏宏.苹果梨果实着色相关PAL基因片段克隆及表达分析[J].延边大学农学学报,2012,34(1):1-5.
猜你喜欢
中国三峡(2023年1期)2023-04-07
基层中医药(2021年2期)2021-07-23
世界最新医学信息文摘(2021年24期)2021-04-29
娃娃乐园·综合智能(2018年5期)2018-04-12
幼儿教育·父母孩子版(2018年10期)2018-03-14
创新作文(小学版)(2017年27期)2017-04-04
今日民族(2016年11期)2016-12-10
云南画报(2015年6期)2015-12-09
大众考古(2015年4期)2015-06-26
食品工业科技(2015年11期)2015-05-05
