
5种桉属植物的m iRNA生物信息学预测
2014-01-17 12:03:22海南医学院生物化学与分子生物学教研室海南海口571199海南大学农学院海南海口570228中国热带农业科学院热带生物技术研究所分析测试中心海南海口571101
安徽农业科学 2014年15期
(1.海南医学院生物化学与分子生物学教研室,海南海口571199;2.海南大学农学院,海南海口570228;3.中国热带农业科学院热带生物技术研究所分析测试中心,海南海口571101)
李崇奇1,2,3,周 鹏2,3* ,蔡望伟1*
桉属(Eucalyptus)植物统称桉树,分类学上属桃金娘科(Myrtaceae),原产于澳大利亚。由于其具有生长速度快、适应性强、材质优良等特点,与松树和杨树并称为世界3大速生树种之一。我国从1890年开始引种栽培桉树,目前栽培面积已达360万hm2,占我国人工林面积的5.8%,占我国木材产量的20%[1]。目前桉树广泛用于房屋建筑、人造板、造纸等领域,同时也是重要的潜在能源植物之一。近年来有研究发现,桉树种植对矿山生态环境的恢复具有积极意义[2]。桉树具有重要的经济价值,也是分子遗传育种改良的重要目标植物之一。识别影响桉树生长速率和其他品质相关的基因对桉树甚至其他林木树种的遗传品质改良具有重要的意义。Lacombe等[3]首次克隆表达了冈尼桉(Eucalyptus gunnii)的木质素合成关键酶肉桂酰CoA还原酶(cinnamyl-CoA reductase,CCR)。Paux 等[4]应用消减杂交技术在冈尼桉中识别224个与木质形成相关的基因,其中1/3的基因与细胞信号转导和细胞壁的合成相关。Ranik和Myburg[5]克隆了6个巨桉(Eucalyptus grandis)全长的纤维素合酶基因,发现纤维素合酶1、2和3主要参与植物次生壁的形成,而纤维素合酶4和5则参与初生壁的合成。Rengel等[6]发现冈尼桉中与木质形成相关的基因主要是与植物初生壁和次生壁形成相关基因及部分转录因子序列。近年来对桉树转录图谱的研究[7-8]更是为从全基因组水平识别和筛选木质相关基因提供了理论基础。然而物种内部很多基因尤其是一些转录因子都受到miRNA的调控,对人类的研究发现30%的基因都受到miRNA的调控[9]。miRNA被认为参与了植物器官的发育、代谢的调节和抗逆反应等一系列复杂生物学过程。笔者拟应用miRtour在线分析工具(http://bio2server.bioinfo.uni-plovdiv.bg/miRTour/)[10]对冈尼桉(Eucalyptus gunnii)、粗皮桉(Eucalyptus pellita)、蓝桉(Eucalyptus globulus)、垂尾桉(Eucalyptus urophylla)和赤桉(Eucalyptus camaldulensis)的表达序列标签(expressed sequence tag,EST)进行分析,然后进行miRNA和靶基因预测,识别与桉树品质相关或疾病相关的miRNA和其调控的靶基因,以期为未来桉树以及其它林木分子遗传育种奠定理论基础。
1 材料与方法
1.1 材料 从美国国家生物技术信息中心(National Center for Biotechnology Information,NCBI)的网站(www.ncbi.nlm.nih.gov)的EST数据库中分别下载冈尼桉、粗皮桉、蓝桉、垂尾桉和赤桉的EST序列,分别为19 841、8 871、28 949、7 440条和58 584条。另外分别从rfam(http://rfam.sanger.ac.uk/)网站和pfam(http://pfam.sanger.ac.uk/)网站下载非编码RNA数据库和蛋白质数据库。
1.2 试验设计 分别将5中桉属植物的EST序列分批递交到miRtour在线界面上传后参数设置如下,与已知miRNA序列能够比对的最小数量(Minimum number of known miRNAs to be aligned)设置为1,miRNA序列与其互补序列的不配对数(Maximum unpaired nt in miR/miR*)设置为6,其他参数默认。下载分析结果可以得到miRNA序列、前体序列、最小自由能、最小自由能指数等相关参数。然后应用blast-2.2.27+软件中的blastn程序将预测到的miRNA前体序列与rfam数据库进行比对,应用blastx程序与pfam数据库进行比对,将evalue参数设置为1e-6,其他参数默认,去除非miRNA序列,即为预测到的miRNA序列前体,相应的miRNA序列即为桉树miRNA序列。将预测到的成熟桉树miRNA序列以fasta格式上传到psrobot网站(http://omicslab.genetics.ac.cn/psRobot/index.php)[11],应用靶基因预测在线工具进行预测,选择巨桉转录组作为对照,参数选择严格模式,同时将分值(score)设置为2.5。
1.3 数据分析 应用bioedit统计miRNA及其前体的序列长度,然后统计miRNA序列每个位点的碱基组成,对其碱基偏倚进行分析。
2 结果与分析
2.1 m iRNA预测 5种桉属所有EST序列经过miRtour分析发现有30条序列具有茎环结构,分别与rfam和pfam数据库比对后发现1条rRNA序列和3条蛋白质序列。共预测到26条miRNA前体序列和19条不同的成熟miRNA序列(表1)。表1中 eca-mir857和 egu-mir857相同,eca-mir164、eglmir164和eur-mir164相同,eca-mir167b和egl-mir167b相同,eca-mir167a和egl-mir167a,但由于在不同物种中发现,所以共命名为24个miRNA。赤桉中发现miRNA序列最多,为12条;而粗皮桉和垂尾桉分别仅发现1条成熟miRNA序。前体序列最小自由能为-19.6~-109.6 kcal/mol,最小自由能指数为0.71~0.94,GC含量为14.0% ~67.9%。
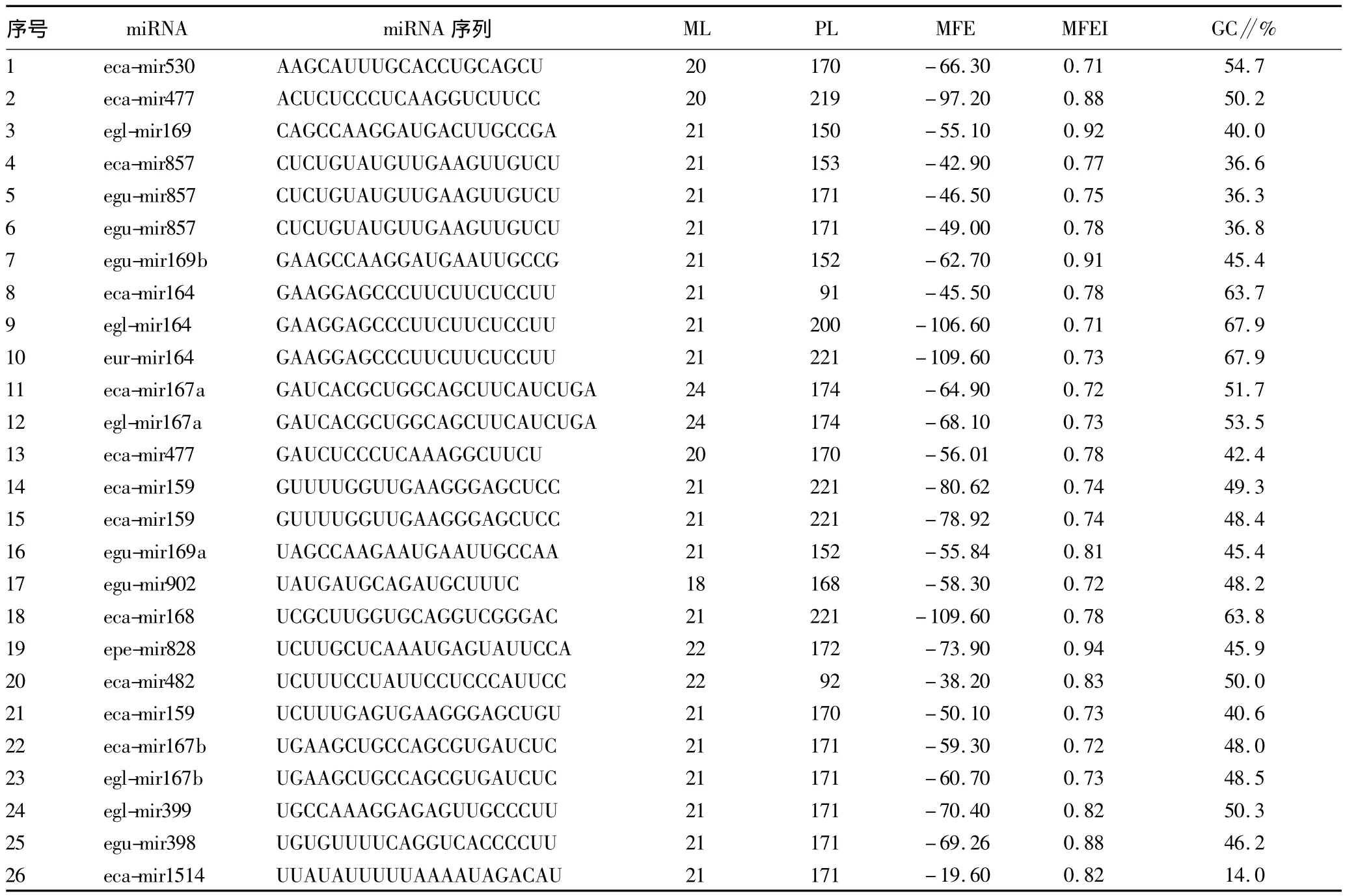
表1 新预测的桉树m iRNA及其前体序列相关参数
2.2 m iRNA和前体的碱基组成特征 预测的miRNA长度为18~24 bp,其中长度为21个碱基的miRNA数量最多,达12个。miRNA序列中嘌呤与嘧啶的比值为1∶1.26,其碱基组成 A∶G∶C∶U 为1∶1.05∶1.15∶1.43,尿嘧啶比例明显偏高。同时发现A、G、C、U出现频率最高的位置分别为5’端第11碱基、第13碱基、第19碱基、第1碱基,其频率分别为52.6%、42.1%、61.1%和52.6%。miRNA前体长度为91~221 bp,平均长度为173 bp,嘌呤与嘧啶的比值为1.00∶1.03,碱基组成 A∶G∶C∶U 为1.00∶1.15∶0.96∶1.25,尿嘧啶的比例最高。
2.3 m iRNA靶基因预测 19个miRNA中有11个预测到了靶基因,共发现104条编码序列受到miRNA调控,去除不能够被注释的序列后共发现87条靶基因序列(表2)。其中32条序列为与疾病抗性相关基因,13条序列为转录因子,16条序列为酶。靶基因数目最多的是mir482,发现40个靶基因。靶基因中与木质形成相关的基因主要为调节MYB转录因子、NAC转录因子和过氧化酶,相应调控木质形成的miRNA主要为mir828、mir902和mir477。
3 结论与讨论
EST序列是从一个随机选择的cDNA克隆进行5’端和3’端单一次测序获得的短的cDNA部分序列,目前被广泛应用于植物miRNA预测研究;尤其是对于缺少基因组信息的物种来说,EST序列是识别miRNA前体序列的重要途径。Zhang等先后应用EST序列分别识别了60个物种的338条miRNA序列[12]和71个物种的481条miRNA 序列[13]。试验经过对桉属5种植物的123 685条序列进行分析,识别了19条不同的miRNA序列,识别效率与多数应用EST序列预测miRNA的研究一致。潘玉欣等对花生(Arachis hypogaea)的研究发现13条miRNA序列[14];郭强等对马铃薯的研究发现22条miRNA序列[15]。然而目前在mirbase数据库(www.mirbase.org)中超过200条成熟microRNA序列的植物已经有13种,miRNA序列最多的蒺藜苜蓿(Medicago truncatula)有756条,因而应用EST识别植物miRNA的效率相对是较低的。这主要可能是由于植物的miRNA序列的初级转录产物很快在DCL等蛋白的作用下加工为为成熟的miRNA序列,从而导致前体序列存在时间较短有关。因而桉树的miRNA还有待于进一步的生物信息学挖掘,或通过芯片技术、基因克隆和小RNA测序等相关实验技术进行识别。
拟南芥和水稻的研究发现其miRNA序列的5’端第1碱基尿嘧啶比例最高[13],而在大豆和亚麻的研究中也发现在第1碱基和第19碱基分别出现频率最高的为尿嘧啶和胞嘧啶[16-17]。试验也发现桉树miRNA序列的5’端第1碱基尿嘧啶出现频率和第19碱基胞嘧啶出现频率都超过了50%。目前认为miRNA 5’端碱基对于其与选择不同Argonaute蛋白结合形成RISC复合物是至关重要的[18]。
桉树是我国木材产量的重要来源之一,筛选与桉树木质形成相关的基因具有重要意义。目前认为影响植物木质形成的转录因子主要包括ARF家族、HD-ZIPIII家族、KAN家族、MYB家族和NAC家族[19];影响木质形成酶主要包括苯丙氨酸裂解酶、4-香豆酸辅酶A连接酶、肉桂醇脱氢酶、过氧化物酶、漆酶和dirigent蛋白[20]。试验结果发现,桉树miRNA靶基因中与木质形成相关的基因主要为调节MYB转录因子、NAC转录因子和过氧化酶,相应参与调控木质形成的miRNA主要为mir828、mir902 和 mir477。Goicoechea等[21]发现桉树MYB2可以结合肉桂酰CoA还原酶基因和肉桂醇脱氢酶基因的启动子区从而调节其转录水平,并在MYB2转基因植物中发现次生细胞壁增厚现象。因而在桉树中miRNA是否为通过影响转录因子的表达水平而影响木质的形成,还是既调节转录因子又同时调节木质形成相关的酶,还需进一步研究。
[1]陈少雄,陈小菲.我国桉树经营的技术问题与思考[J].桉树科技,2013,30(3):52-59.
[2]时彧,罗杰.桉树对矿山环境的改善[J].安徽农业科学,2014,42(2):483-486.
[3]LACOMBE E,HAWKINS S,VAN DOORSSELAERE J,et al.Cinnamoyl CoA reductase,the first committed enzymeof the lignin branch biosynthetic pathway:cloning,expression and phylogenetic relationships[J].Plant J,1997,11(3):429-441.
[4]PAUX E,TAMASLOUKHTM,LADOUCEN S,etal.Identification of genes preferentially expressed during wood formation in Eucalyptus[J].Plant Mol Biol,2004,55(2):263-280.
[5]RANIK M,MYBURG A A.Six new cellulose synthase genes from Eucalyptus are associated with primary and secondary cell wall biosynthesis[J].Tree Physiol,2006,26(5):545-556.
[6]RENGEL D,SAN CLEMENTE H,SERVANT F,et al.A new genomic resource dedicated to wood formation in Eucalyptus[J].BMC Plant Biol,2009,27;9:36.
[7]HENDRE P S,KAMALAKANNAN R,VARGHESE M.High-throughput and parallel SNP discovery in selected candidate genes in Eucalyptus camaldulensis using Illumina NGS platform[J].Plant Biotechnol J,2012,10(6):646-656.
[8]MIZRACHIE,HEFERCA,RANIKM,etal.De novo assembled expressed gene catalog of a fast-growing Eucalyptus tree produced by IlluminamRNA-Seq[J].BMCGenomics,2010,11:681.
[9]LEWISB P,BURGE C B,BARTEL D P.Conserved seed pairing,often flanked by adenosines,indicates that thousands of human genes are microRNA targets[J].Cell,2005,120(1):15-20.
[10]MILEV I,YAHUBYAN G,MINKOV I,et al.miRTour:PlantmiRNA and target prediction tool[J].Bioinformation,2011,6(6):248-249.
[11]WU H J,MA Y K,CHEN T,etal.PsRobot:aweb-based plant small RNA meta-analysis toolbox[J].Nucleic Acids Research,2012,40(W1):22-28.
[12]ZHANGBH,PANX P,WANGQ L,etal.Identification and characterization of new plantmicroRNAs using EST analysis[J].Cell Res,2005,15(5):336-360.
[13]ZHANG B,PAN X,CANNON C,et al.Conservation and divergence of plantmicroRNA genes[J].Plant J,2006,46(2):243-259.
[14]潘玉欣,刘恒蔚.花生miRNA与其靶基因的生物信息学预测[J].中国油料作物学报,2010,32(2):290-294.
[15]郭强,项安玲,杨清,等.利用 EST及生物信息学方法挖掘马铃薯中miRNA 及其靶基因[J].科学通报,2007,52(14):1656-1664.
[16]ZHANGB,PANX,STELLWAGE J.Identification of soybeanmicroRNAs and their targets[J].Planta,2008,229(1):161-182.
[17]NEUTELINGS G,FÉNART S,LUCAU-DANILA A,et al.Identification and characterization ofmiRNAs and their potential targets in flax[J].J Plant Physiol,2012,169(17):1754-1766.
[18]THIEME C J,SCHUDOMA C,MAY P,etal.Give It AGO:The Search for miRNA-Argonaute Sorting Signals in Arabidopsis thaliana Indicates a Relevance of Sequence Positions Other than the 5'-Position Alone[J].Front Plant Sci,2012,3:272.
[19]DEMURA T,FUKUDA H.Transcriptional regulation in wood formation[J].Trends Plant Sci,2007,12(2):64-70.
[20]石海燕,张玉星.木质素生物合成途径中关键酶基因的分子特征[J].中国农学通报,2011,27(5):288-291.
[21]GOICOECHEA M,LACOMBE E,LEGAY S,et al.EgMYB2,a new transcriptional activator from Eucalyptus xylem,regulates secondary cell wall formation and lignin biosynthesis[J].The Plant Journal,2005,43(4):553.
猜你喜欢
科学大众·小诺贝尔(低幼)(2024年5期)2024-12-31 00:00:00
天津医科大学学报(2021年1期)2021-12-05 11:11:05
教学考试(高考生物)(2020年6期)2020-11-23 05:25:56
食品与生物技术学报(2020年8期)2020-01-06 08:00:56
科学24小时(2019年5期)2019-06-11 08:39:38
发明与创新(2019年9期)2019-03-26 02:22:48
环球时报(2019-01-03)2019-01-03 09:06:44
现代检验医学杂志(2016年5期)2016-08-20 03:17:08
广西林业科学(2016年2期)2016-03-20 05:53:22
广西林业科学(2016年4期)2016-03-16 05:44:51
