
速生树种尾巨桉和竹柳幼苗的光合特性和根系特征比较
2014-01-04 09:47李吉跃
中南林业科技大学学报 2014年1期
邱 权,潘 昕,李吉跃,何 茜,苏 艳,董 蕾
(华南农业大学 林学院,广东 广州 510642)
速生树种尾巨桉和竹柳幼苗的光合特性和根系特征比较
邱 权,潘 昕,李吉跃,何 茜,苏 艳,董 蕾
(华南农业大学 林学院,广东 广州 510642)
选择华南地区两种速生树种尾巨桉Eucalyptus urophylla×Eucalyptus grandis和竹柳Salixspp.幼苗,通过测定其各自光响应曲线特征、光合生理指标(净光合速率Pn、气孔导度Gs、胞间CO2浓度Ci、蒸腾速率Tr和水分利用效率WUE)日变化规律以及根系特征指标(根长、表面积、体积和根尖数),对比此两种树种光合生长能力、水分利用和消耗能力以及根系吸收水分和养分能力的差异。结果表明:(1)净光合速率、蒸腾速率日变化规律呈明显的“双峰”曲线,峰值均出现在10:00和16:00,而水分利用效率日变化规律不明显,尾巨桉和竹柳白天瞬时WUE变化范围分别介于1.863~3.941 μmol/mmol和2.645~3.894 μmol/mmol之间。(2)Pn与温度和光照强度呈显著正相关,Gs与空气相对湿度呈显著正相关,Tr与温度和空气相对湿度均呈显著正相关,而Ci和WUE与环境因子相关性均不显著。(3)竹柳潜在最大净光合速率以及白天不同时间瞬时净光合速率均高于尾巨桉,显示出了更优的潜在和实际光合生长能力,而尾巨桉全根系和细根(直径≤2 mm)的长度、表面积、体积和根尖数均高于竹柳,表明尾巨桉具有更强的吸水和吸肥能力。(4)综合来看,竹柳光合生长能力强,水分利用效率高,但叶片水分消耗较大,而尾巨桉节水性好,吸水吸肥能力更强,两个速生树种各有优劣。
尾巨桉;竹柳;光合特性;根系特征;日变化
光合作用和蒸腾作用是植物重要的生命活动,其中光合作用是植物生长的生理基础,蒸腾作用耗水则是树木水分散失的主要途径,而水分利用效率则是评价植物水分消耗和利用能力的主要评价指标。对于植物光合特性的研究,近几年来,国内外学者主要采用的是Li-6400光合系统测定方法。其中关于光合作用和蒸腾作用的季节动态变化、日动态变化以及光响应曲线等方面的内容研究较多[1-7],研究方法比较成熟。根系是植物吸收水分和养分的主要器官,其中细根(直径≤2 mm)是主要的吸收和代谢根,粗根主要起过渡和贮藏水分和养分的作用,虽然细根生物量占根系总生物量比例低于30%,但其根系特征值占全根比例较大[8-10],因此细根特征值对于评价植物对水分和养分的吸收能力尤其重要。尾巨桉Eucalyptus urophylla×Eucalyptus grandis是尾叶桉E. urophylla和巨桉E. grandis杂交种,为华南地区主要的桉树品种,因此受到了较多的关注[9]。竹柳又称美国竹柳,为杨柳科Saliaceae柳属Salix乔本植物,美国寒竹、朝鲜柳、筐柳组合杂交选育的优良杂交品系,近几年从美国引入国内栽种,由于其生长快、耐盐碱、耐水淹、材质优良、干形优美,很快成为了引起普遍关注的新兴起速生树种[12-13]。尾巨桉和竹柳作为华南地区的两种重要速生树种,水分消耗和利用问题一直是围绕速生树种争议较多的热点,而关于两者光合生长能力、水分消耗和利用能力的比较鲜见报道,针对其与水分和养分吸收能力紧密相关的根系特征的对比研究也较少。
本研究主要采用Li-6400光合系统测定方法研究尾巨桉和竹柳幼苗的光响应曲线特征、瞬时净光合速率、蒸腾速率和水分利用效率的日变化规律,并通过根系扫描和图像软件分析的方法测定苗木根系分级特征值(根系长度、表面积、体积和根尖数),综合对比此两种苗木光合生长能力、水分消耗和利用能力以及水分和养分吸收能力,分析两者的共性和差异性,并为华南地区速生树种综合性能评价提供一些理论参考。
1 材料与方法
1.1 试验材料
选择常见桉树品系尾巨桉(广林-9号)无性系1年生组培苗和竹柳1年生扦插苗,2011年4月初将所有供试材料上盆,每盆1株,每种苗木各上盆30株。所用花盆规格为200 mm×150 mm,基质土壤取自华南农业大学树木园,其田间持水量为(26.87±2.07)%,容重为(1.34±0.07)g/cm3。定期浇水,除草,防病虫害,保证苗木正常生长。实验测定开始时苗木基本情况见表1。

表1 苗木生长情况Table 1 Growth status of seedlings
1.2 研究方法
1.2.1 光响应曲线
20011年7月初,用Li-6400便携式光合作用分析系统(美国)于上午9:00~11:30进行测定。测定过程使用红蓝光源,将叶室温度设定为稍高于空气温度,光强设置为0、20、50、100、150、200、400、600、800、1 000、1 200、1 500、1 800 μmol·m-2s-1。每种苗木选择 3 株,每株测定功能叶3片。
1.2.2 气体交换参数日变化
2011年7月初,用Li-6400便携式光合作用分析系统(美国)选择典型晴天于8:00、10:00、12:00、14:00、16:00和 18:00,每间隔 2h测定一次叶片瞬时净光合速率(Pn),蒸腾速率(Tr),而水分利用效率用公式:水分利用效率(WUE)=净光合速率/蒸腾速率求出。仪器同时记录气孔导度(Gs)、胞间 CO2浓度(Ci)、环境间 CO2浓度(Ca)、环境温度(T) 、光照强度(PAR)和相对湿度(RH)。每种苗木选择3株,每株测定3片功能叶,观察苗木白天不同时间的气体交换参数变化规律。
1.2.3 根系特征
于2011年7月初,每种苗木分别随机选取5株苗木的完整根系(外业获取的根系要放入冰盒中保鲜,防止根系脱水,体积变小)置于密封袋带回实验室后用根系扫描系统分析根系形态特征(总根长、表面积、体积、根尖数、分叉数),并对根系进行分级,重点研究微细根(d≤2 mm)根长、表面积、体积及根尖数。根系分析方法参考王冉等[14]和王力朋等[15],根系划分为细根(f i ne roots, 直径≤2 mm)和粗根(coarse roots,>2 mm)。根系扫描仪为中晶ScanMaker i800 Plus,分析软件为万深LA-S植物根系分析系统(杭州万深检测科技有限公司)。处理前,将根系样品从冰盒中取出,用清水小心冲洗掉根系表面的泥土。如遇到很细的根,则在根系下面放置100目筛以防止脱落的根系被水冲走。
1.3 数据处理与分析
用Excel作图和SPSS19.0对试验数据进行方差分析、多重比较、相关性分析。
2 结果与分析
2.1 苗木的光响应曲线特征比较
由图1可以看出,尾巨桉和竹柳苗木的净光合速率随光照强度总体变化趋势相同,但具体变化规律有所不同,需要进行光响应曲线模型的拟合进一步对比相关光合生理参数。本研究采用拟合效果较好的非直角双曲线(Farquhar模型)[16],利用SPSS统计软件中的非线性回归方法来估计模型参数[17],从而得出两种苗木光响应曲线各特征参数值(见表2)。光响应曲线(即Pn-PAR曲线)描述的是植物净光合速率Pn与光合有效辐射PAR之间的关系,可以提供植物光合特性的相关生理参数,是判定植物光合效率的重要手段。光补偿点LCP(light compensation point)和光饱和点 LSP(light saturation point)是分别反映植物利用弱光和强光能力的评价指标,而最大净光合速率(Pnmax)是反映植物潜在生长能力的重要指标。竹柳的最大净光合速率为 14.66 μmol·m-2s-1,高于尾巨桉(12.19 μmol·m-2s-1),可见竹柳潜在光合生长能力更强。通过比较两种苗木的光补偿点和光饱和点可知,竹柳的光补偿点(17.61 μmol·m-2s-1)和光饱和点(956.00 μmol·m-2s-1)均明显大于尾巨桉,可见,光补偿点和光饱和点均较高的竹柳更加喜光。总体来讲,竹柳可利用光的范围较大,可见其具有较高的光合能力。
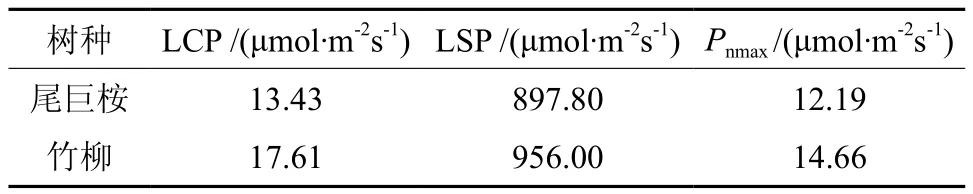
表2 光响应曲线的特征参数值Table 2 Characteristic parameters of photo response curves
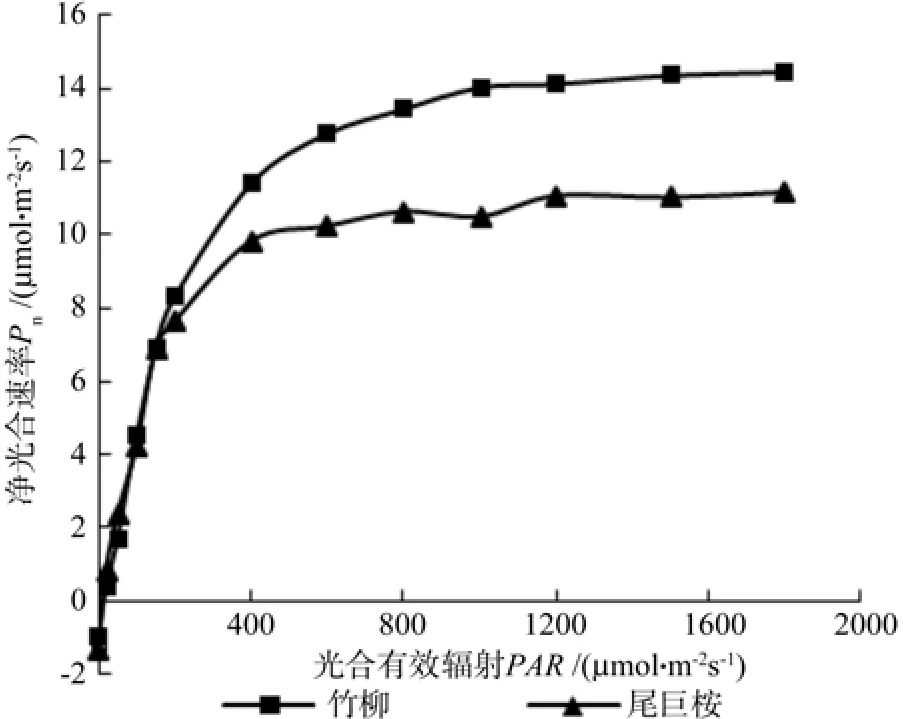
图1 苗木光响应曲线Fig. 1 Light response curves of seedlings
2.2 净光合速率Pn日变化特征
一般认为,当土壤水分充足时,太阳光照是植物光合作用主要的限制因素,净光合速率会随着光照强度的变化随之改变,呈现出日动态变化特征。但当中午光照强度较大时,相对湿度小,水分供应紧张,植物会出现“午休”现象,净光合速率迅速下降。由图2可知,尾巨桉和竹柳苗木的净光合速率日变化呈典型的“双峰”曲线,峰值均出现在10:00和16:00,但10:00时两种苗木的净光合速率均高于16:00。其中竹柳苗木净光合速率第一个峰值为(14.966±0.277)μmol·m-2s-1,高于尾巨桉(11.753±0.539)μmol·m-2s-1。第两个峰值也是竹柳(12.074±0.868)μmol·m-2s-1>尾巨桉(9.322±0.494)μmol·m-2s-1。就变化趋势而言,尾巨桉和竹柳苗木有所不同。从图1中日变化曲线可以看出,上午10:00以后,两种苗木净光合速率均呈下降趋势,拐点均出现在12:00,可见两种苗木虽然存在“午休”现象,但尾巨桉午休的时间更长,在12:00~14:00时间段内净光合速率一直较低,到14:00以后其光合作用才慢慢恢复到正常状态,说明尾巨桉对水分更为敏感,当温度很高,空气湿度较低时(见图3),由于植物需要控制水分的蒸散,所以长时间关闭了部分气孔,进而导致光合速率也随之下降。对比两种苗木不同时间光合速率可知,竹柳均明显高于尾巨桉,表现出了更强的光合生长能力。
2.3 蒸腾速率Tr日变化特征
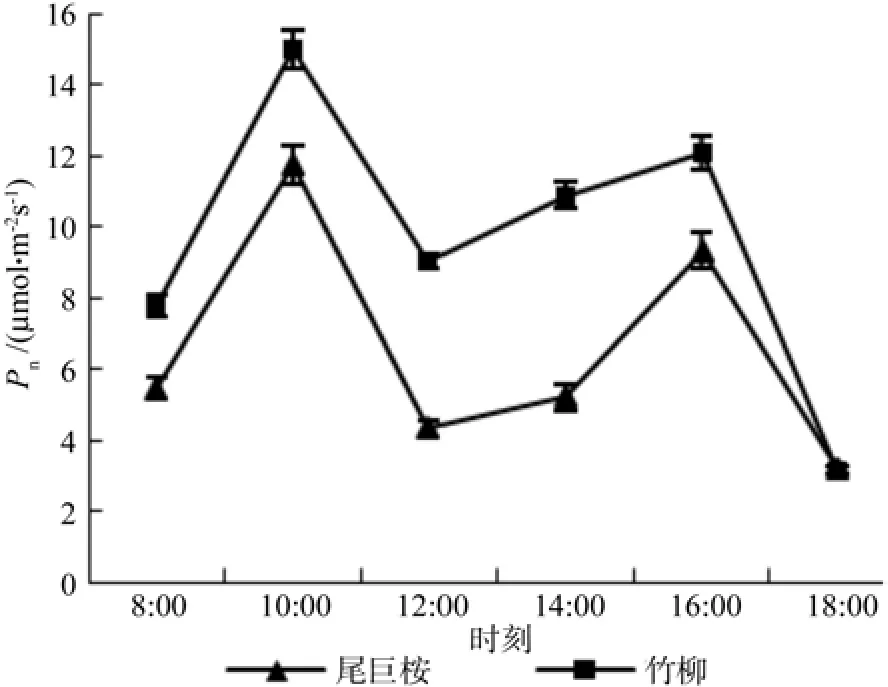
图2 苗木白天不同时间净光合速率(Pn)变化Fig.2 Daily changes of net photosynthesis rate Pn of seedlings
由图3可知,两种苗木蒸腾速率日变化趋势总体上与净光合速率一致,也呈典型的“双峰”曲线,两个峰值分别出现在10:00和16:00。对比两种苗木两个峰值发现,竹柳两个净光合速率峰值均高于竹柳,其中竹柳10:00和16:00的两个净光合速率峰值分别为(4.477±0.201)mmol·m-2s-1和(4.054±0.311)mmol·m-2s-1,而尾巨桉10:00和16:00的两个净光合速率峰值分别为(4.155±0.215)mmol·m-2s-1和(3.852±0.377)mmol·m-2s-1。从变化曲线上看,两种苗木在12:00~14:00之间保持了较长时间的低蒸腾速率,主要原因是也是此时间环境温度最高,空气相对湿度最低,诱使叶片部分气孔关闭,而气孔关闭是植物蒸腾下降的重要因素[18],从而可以减少水分的蒸发。从图4可以看出,尾巨桉每一个时期的蒸腾速率均明显低于竹柳,可见在叶片水平上来看,尾巨桉的水分消耗更少,显示出了更强的节水能力。
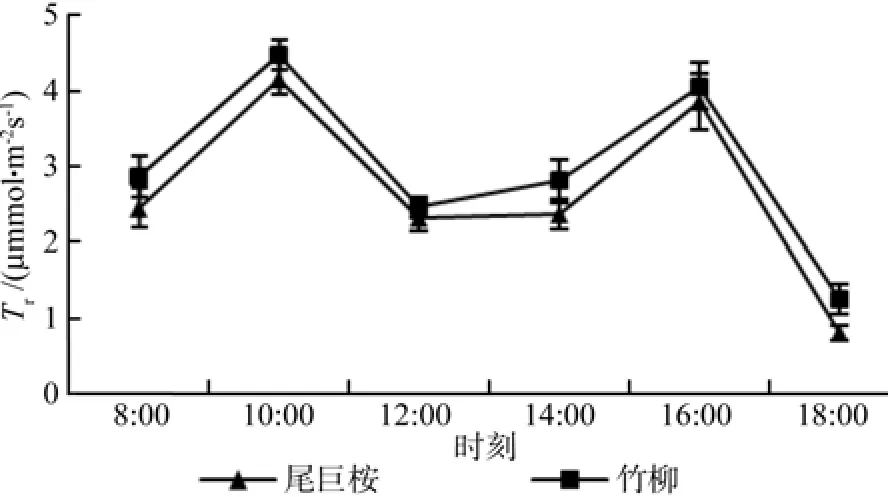
图3 苗木白天不同时间蒸腾速率(Tr)变化Fig.3 Daily changes of transpiration rate Tr of seedlings
2.4 气孔导度Gs和胞间CO2浓度Ci日变化特征
气孔是叶片控制光合和蒸腾的重要因素,其开合和关闭主要受环境因子光照、温度和湿度等因素影响,而胞间CO2浓度则能一定程度反应植物叶片光合作用的状况。图4和图5可以看出,两种苗木气孔导度和胞间CO2呈现出不同的日变化特征。其中,两种苗木气孔导度呈现出双峰曲线,双峰均出现在10:00和16:00,但尾巨桉最低气孔导度出现在12:00,而竹柳出现在14:00。胞间CO2浓度呈现出总体下降的趋势,8:00时,苗木光合作用才开始,细胞内CO2浓度还未进行大量消耗,而随着日进程的变化,其光合速率开始上升,需要消耗大量的CO2,所以细胞内CO2慢慢开始下降。当进入12:00时,尾巨桉气孔逐渐关闭,气孔导度最低,光合速率也下降至最低值,而竹柳则在14:00出现气孔大量关闭的情况,从而细胞内CO2的消耗进程得到缓解。14:00以后,随着气孔导度的增加,光合作用的慢慢恢复,细胞内开始积累CO2,以保障光合作用对CO2的大量需求。
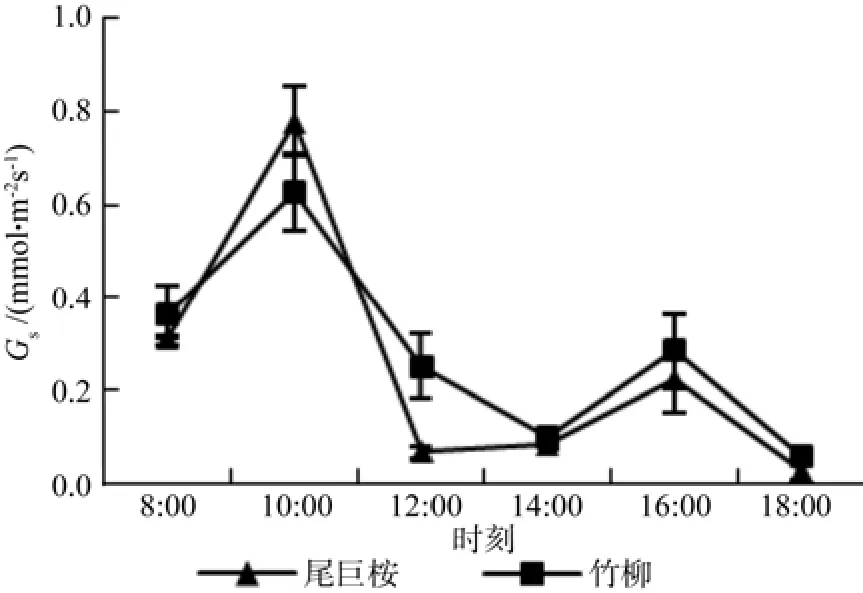
图4 苗木白天不同时间气孔导度(Gs)变化Fig.4 Daily changes of Gs of seedlings
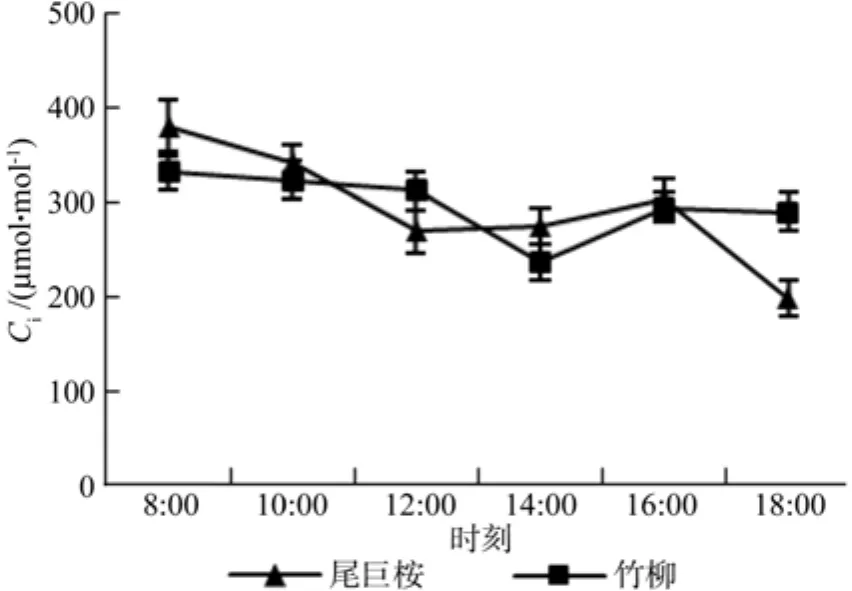
图5 苗木白天不同时间胞间CO2浓度(Ci)变化Fig.5 Daily changes of Ci of seedlings
2.5 瞬时水分利用效率(RWUE)日变化
由图6可以看出,两种苗木叶片瞬时RWUE呈现出不同的日变化规律,尾巨桉呈单峰曲线,而竹柳变化趋势无明显规律。其中尾巨桉和竹柳白天瞬时RWUE变化范围分别介于1.863~3.941 μmol/mmol和 2.645~ 3.894 μmol/mmol之间。对比两种苗木在每个时间段的叶片WUE发现,白天大部分时间(8:00~16:00)竹柳的叶片瞬时水分利用效率高于尾巨桉,仅18:00时出现竹柳低于尾巨桉的现象,因此可以说明,在叶片水平上竹柳的水分利用效率总体高于尾巨桉,显示出了更高效的水分利用能力。
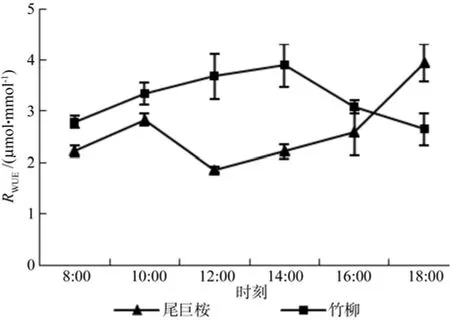
图6 苗木白天不同时间瞬时水分利用效率RWUE变化Fig.6 Daily changes of RWUE of seedlings
2.6 光合指标与环境因子相关性
光合参数的日变化特征主要受自身结构和环境因子温度、相对湿度、大气CO2浓度和光照强度等因素影响[19-20]。从图7可以看出,温度和光照强度呈现出明显的单峰曲线,峰值均出现在12:00,相对湿度则呈现出“U”型变化趋势,12:00空气相对湿度最低,而大气CO2浓度则呈逐渐下降趋势。需要注意的是,中午12:00时出现的高温、低湿、高光强可能是植物午休和气孔关闭的主要原因。为更好地分析光合生理参数与环境因子的关系,对光合生理指标和环境因子进行相关性分析(见表3),结果表明净光合速率与温度和光照强度呈显著正相关,气孔导度与空气相对湿度呈显著正相关,蒸腾速率与温度和空气相对湿度均呈显著正相关,而胞间CO2浓度和WUE与环境因子相关性均不显著。此外,净光合速率、蒸腾速率和气孔导度这3个光合生理指标之间存在显著正相关关系,表现出一定的相互促进关系。
2.7 根系特征比较
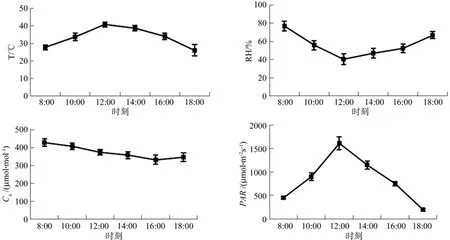
图7 白天不同时间环境因子变化Fig.7 Daily changes of environmental factors
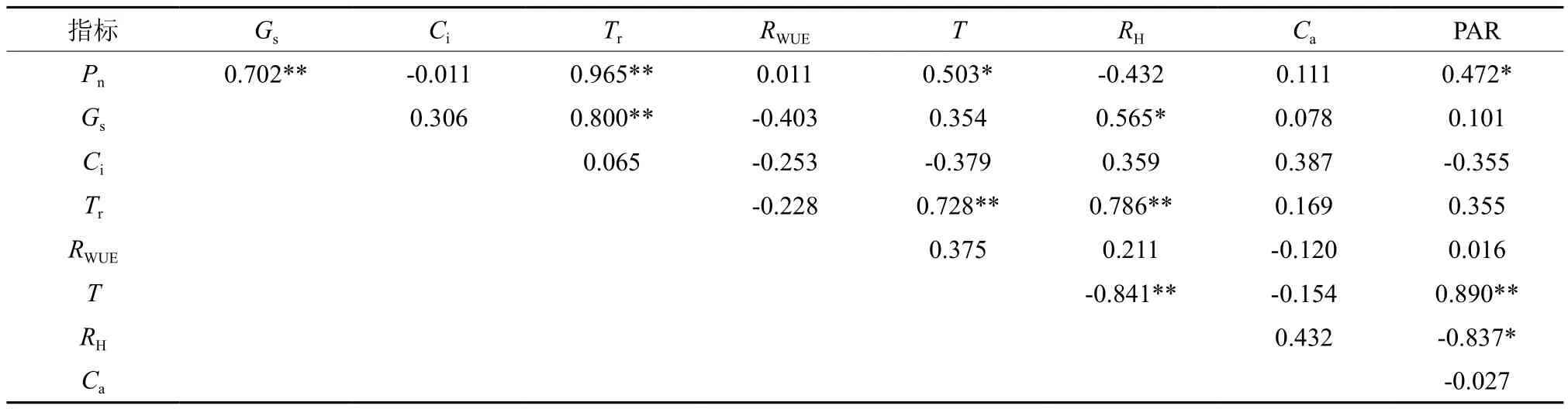
表3 光合生理指标与环境因子的相关性†Table 3 Correlation of photosynthetic indexes and environmental factors
由图8、图9、图10、图11可以看出,尾巨桉全根总长度、表面积、体积和根尖数均明显高于竹柳,并且尾巨桉和竹柳苗木细根(直径≤2 mm)总根长、表面积、体积和根尖数占全根对应特征值的比例均明显高于粗根。其中尾巨桉苗木总根长、表面积、体积和根尖数占全根对应特征值的比例分别为87.88%、90.79%、66.69%和85.99%,而竹柳苗木总根长、表面积、体积和根尖数占全根对应特征值的比例分别为89.96%、84.48%、55.24%和63.80%,可见细根特征值所占比例较大,可以表征大部分的根系特征信息。因此,需要对两种苗木的细根特征值进行全面对比。
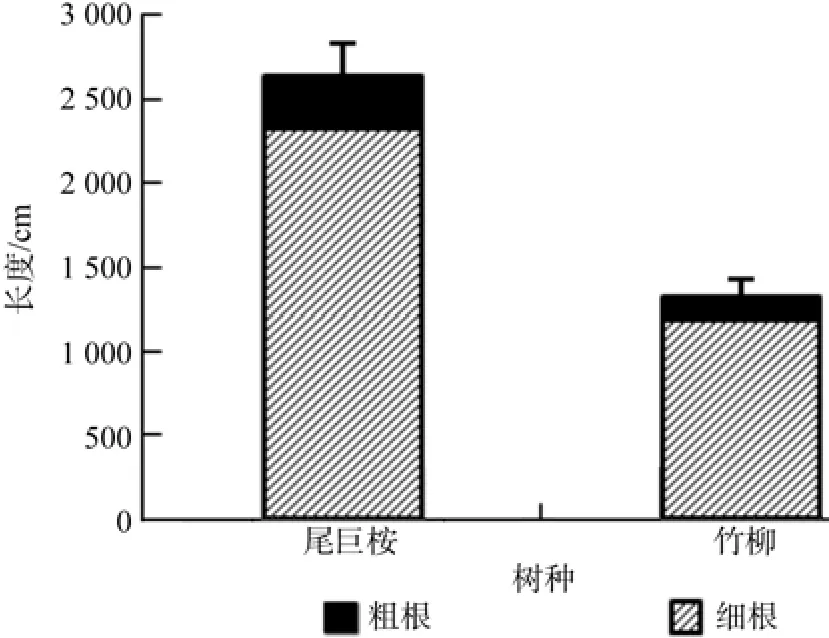
图8 苗木根系长度对比Fig.8 Length of root system
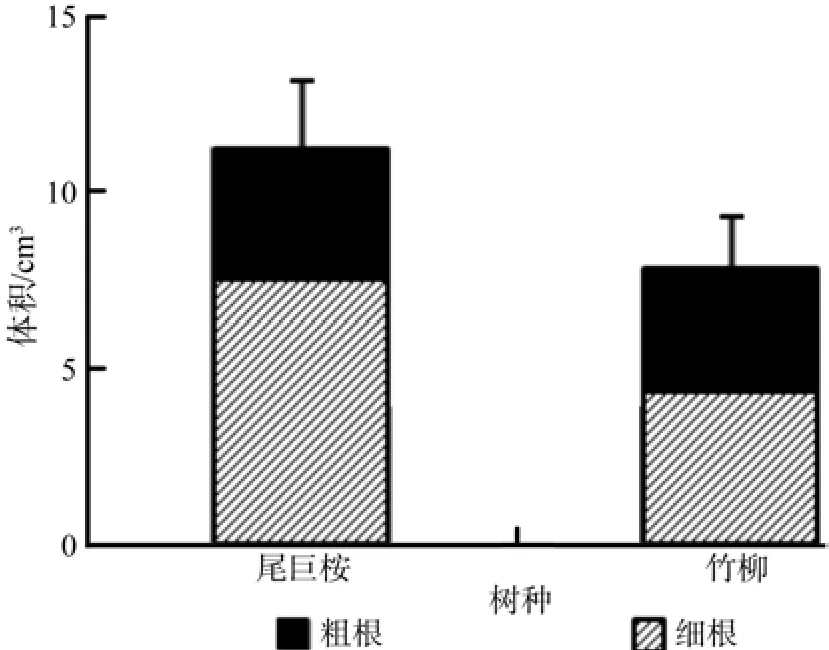
图10 苗木根系体积对比Fig.10 Comparison of root system volume of two trees
3 结论与讨论
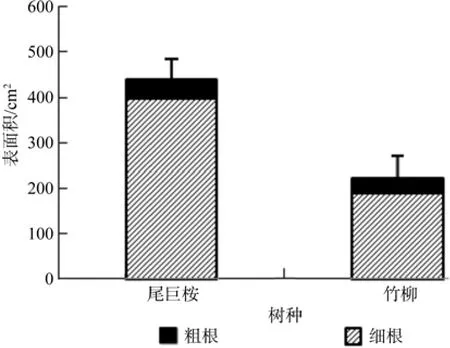
图9 苗木根系表面积对比Fig.9 Surface area of root system
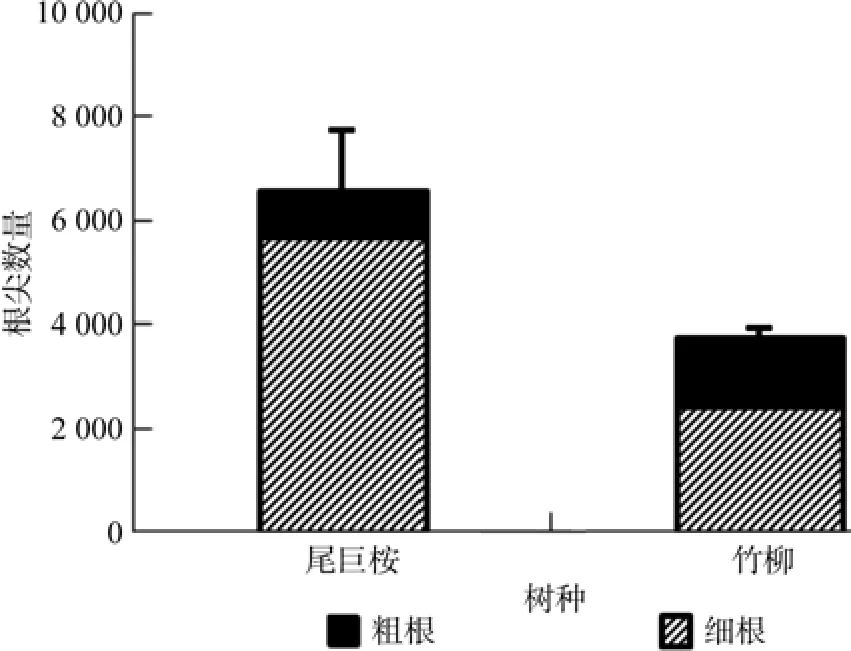
图11 苗木根尖数对比Fig.11 Comparison of numbers of root tips of two trees
有研究表明,植物光合生理指标与环境因子具有显著相关性[2-3,21-22]。本研究中苗木叶片净光合速率与温度和光照强度呈显著正相关,光照是光合作用的来源,而温度主要表现为对酶活性和膜透性的影响。气孔导度与空气相对湿度呈显著正相关,表明空气湿度对气孔关闭和开合影响最为显著。蒸腾速率与温度和空气相对湿度均呈显著正相关,主要原因是一定程度的升温会加快叶研究结果表明尾巨桉细根总长度、表面积、体积和根尖数分别为2 321.58 cm、399.72 cm2、7.48 cm3和5 642,分别比竹柳高95.94%、110.10%、73.56%和137.66%。一般认为,根系越长说明垂直扎根深或者侧根水平伸展范围广,植物根系可以吸收土层深度较深或水平方向上距离较远的土壤水分和养分,根表面积和体积越大表明可吸收利用水分和养分范围越大,根尖数越多说明须根越多,吸水吸肥能力越强,因此可以说明尾巨桉的根系特征显示出了更强的水分和养分吸收能力。片蒸腾,而空气相对湿度较低时,植物便会降低蒸腾来保存水分,因此温度和空气相对湿度均是正向显著影响植物蒸腾速率。大量研究表明,随着环境因子的改变,植物光合作用和蒸腾作用会呈现出明显的日变化规律[2-3,5,23]。本研究中光合速率呈明显的“双峰”曲线,与已经报道的对于华南地区阔叶树种楝叶吴茱萸Evodia glabrifolia、石斑木Rhaphiolepis indica和任豆Zenia insignis的研究结论一致[23],并且尾巨桉和竹柳白天净光合速率峰值明显高于此3种阔叶树种,说明速生树种光合生长能力相对优于一般阔叶树种。另据赵曦阳等[1]对楸树Catalpa bungei无性系光合特性的研究显示楸树净光合速率、蒸腾速率、胞间CO2浓度和气孔导度均呈现出明显的日变化趋势,在本研究中进一步证实了此结论。另一方面,微细根是主要的吸收和代谢根,表皮薄、幼嫩、膜透性强,具有吸收表面积大、生理活性强等特点,是树木水分和养分吸收的主要器官[8,24]。本研究中两种速生树种细根特征值占全根的比例较大,其中两种苗木细根长和表面积占全根的比例甚至超过了80%,可见速生树种在苗期生长旺盛季节主要依赖于其细根的强大吸收能力来获取大量水分和养分,以满足植物快速生长时的大量需求。
从光响应曲线特征和净光合速率日变化特征综合来看,竹柳潜在最大净光合速率以及白天光合生长能力均高于尾巨桉,表明竹柳的潜在和实际光合生长能力均优于尾巨桉。根系特征方面,尾巨桉根系特征值尤其是细根(直径≤2 mm)的长度、表面积、体积和根尖数均高于竹柳,表明尾巨桉具有更强的吸水和吸肥能力。此两种树种均为华南地区速生树种,从研究结果来看,可能两个树种代表着两种不同的速生策略。其中竹柳主要通过叶片高光合来提高植株生产力,而尾巨桉则主要通过强大的根系分布来提高其获取水分和养分的能力。从叶片蒸腾速率来看,竹柳的高蒸腾速率可能引起更多的水分消耗,因此在叶片水平来讲尾巨桉节水性相对较好。水分利用效率方面,研究结果表明竹柳总体上显示出了比尾巨桉更高效的水分利用效率。综合来看,竹柳光合生长能力强,水分利用效率高,但水分消耗较大,而尾巨桉节水性好,吸水吸肥能力更强,两个速生树种各有优劣,因此在不同地区造林树种选择时需要全面考量其各自综合性能。
[1] 王旭军,潘百红,程 勇,等.红榉不同种源光合特性的比较[J].中南林业科技大学学报,2013,33(6):37-42.
[2] 蒋 燚,李志辉,朱积余,等.不同红锥种源幼林光合特性日变化测定与分析[J]. 中南林业科技大学学报,2013,33(6):43-47.
[3] 赵曦阳,王军辉,张金凤,等.楸树无性系光合特性研究[J].西北植物学报,2011,31(8):1665-1671.
[4] 伍维模,李志军,罗青红,等.土壤水分胁迫对胡杨灰叶胡杨光合作用-光响应特性的影响[J].林业科学,2007,43(5):30-35.
[5] 郭二果,马颖聪,常金宝.伊金霍洛旗沙柳幼苗光合、蒸腾强度日动态变化[J].内蒙古科技与经济, 2005,(11):103-106.
[6] Wang M C,Wang J X,Shi Q H,et al.Photosynthesis and water use eff i ciency of platycladus orientalis and robinia pseudoacacia saplings under steady soil water stress during different stages of their annual growth period[J].Journal of Integrative Plant Biology,2007,49(10):1470-1477.
[7] 黄志宏,田大伦,闫文德,等.城市樟树人工林冠层光合作用的时空特征[J].中南林业科技大学学报,2011,31(1):38-46.
[8] Usman S,Singh S P,Rawat Y S,et al.Fine root decomposition and nitrogen miner ralisation patterns inQuercus leucotrichophoraandPinus roxburghiiforest in central Himalaya[J].Forest Ecology and Management,2000,131(13):191-199.
[9] Cheng S, Widden P, Missier C. Light and tree size influence belowground development in yellow birch and sugar maple[J].Plant and Soil, 2005,270(1):321-330.
[10] 李林锋,刘新田.干旱胁迫对桉树幼苗的生长和某些生理生态特性的影响[J]. 西北林学院学报,2004,19(1):14-17.
[11] 侯元兆.科学地认识我国南方发展桉树速生丰产林问题[J].世界林业研究,2006,19(3):71-76.
[12] 吕德文.竹柳引种试验初报[J].防护林科技,2011(5):42-43,53.
[13] 王文成,郭艳超,李克晔,等.盐胁迫对竹柳种苗形态及生理指标的影响[J].华北农学报,2011,26(增刊):143-146.
[14] Farquhar G D, Oleary M H, Berry J A. On the relationship between carbon isotope discrimination and the intercellular carbon dioxide concentration in leaves[J].Australian Journal of Plant Physiology,1982,9(11):121-137.
[15] 刘宇锋,萧浪涛,童建华,等.非直线双曲线模型在光合响应曲线数据分析中的应用[J].农业基础科学,2005,21(8):76-79.
[16] 王 冉,李吉跃,张方秋,等.不同施肥方法对马来沉香和土沉香苗期根系生长的影响[J].生态学报,2011,31(1):908-0106.
[17] 王力朋,晏紫伊,李吉跃,等.指数施肥对楸树无性系生物量分配和根系形态的影响[J].生态学报,2012,32(23):7452-7462.
[18] 黄颜梅,张 健,罗成德.树木抗旱性研究[J].四川农业大学学报,1997,15(1) : 49-54.
[19] Calfapietra C, Tulva I, Eensalu E,et al.Canopy profiles of photosynthetic parameters under elevated CO2and N fertilization in a poplar plantation[J] . Environmental Pollution, 2005, 137(3): 525- 535.
[20] Li S W, Pezeshki S R, Goodwin S. Effects of soil moisture regimes on photosynthesis and growth in cattail (Typha latifolia) [J]. Acta Oecologica, 2004, 25(1-2): 17-22.
[21] 侯晓改.4个牡丹品种光合特性的比较研究[J].河南农业大学学报,2007,41(5):527-530.
[22] 杨江山,常永义,种培芳,等.3个樱桃品种光合特性比较研究[J].园艺学报,2005,32(5):773-777.
[23] 邱 权,潘 昕,何 茜,等.华南地区3种苗木生长旺盛时期光合特性及蒸腾耗水日变化规律的比较[J].华南农业大学学报,2012,33(4):524-528,549.
[24] 卫 星,张国珍. 树木细根主要研究领域及展望[J].中国农学通报,2008,24(5):143-147.
Comparison on photosynthetic and roots characteristics of Eucalyptus urophylla × Eucalyptus grandis and bamboo-willow seedlings
QIU Quan, PAN Xin, LI Ji-yue, HE Qian, SU Yan, DONG Lei
(College of Forestry, South China Agricultural University, Guangzhou 510642, Guangdong, China)
Two fast-growing trees (Eucalyptus urophylla×Eucalyptus grandis) andSalixspp. seedlings were selected as the tested materials, theirs differences in ability of photosynthesis, water use and consumption and root absorption of water and nutrients were compared after measuring the light response curve characteristics, daily change of photosynthetic indexes [net photosynthetic rate(Pn), stomatal conductance(Gs), intercellular CO2concentration(Ci), transpiration rate (Tr) and water use efficiency (RWUE) ]and the characteristics values length, surface area, volume and amount of root tips of root system were measured. The results show that (1)the daily changing trends ofPnandTrof two kinds of seedlings were “twink peak”curve, and their peaks both appeared at a.m.10:00 and p.m.16:00, but the daily changing rule ofRWUEof the seedlings was not obvious; (2)Pnshowed a signif i cant positive correlation with the temperature and light intensity,Gshad a signif i cant positive correlation with air relative humidity, andTrshowed a signif i cant positive correlation with temperature and air relative humidity, whereasCiandRWUEshowed no signif i cant correlation with the environmental factors; (3) Potential maximumPn.m and instantaneousPn, I during daytime of bamboo-willow was higher than that ofE. urophylla×E. grandis, so it showed that the potential and actual growth ability through photosynthesis of bamboo-willow was superior to that ofE.urophylla×E. grandis, on the other hand, the length, surface area, volume and amount of root tips of root system especially fi ne root(diameter ≤ 2 mm ) ofE. urophylla×E. grandiswere higher than that of bamboo-willow, thereforeE. urophylla×E. grandisshowed better ability of water and nutrients absorption; (4) in general, bamboo-willow showed superior growth ability and higherRWUEthanE.urophylla×E. grandis, but its water consumption of leaves was larger, in contrast,E. urophylla×E. grandishad signif i cant advantage in water saving and its ability of water and nutrients absorption was stronger, thus the two fast-growing tree species each had advantages and disadvantages.
Eucalyptus urophylla×Eucalyptus grandis;Salixspp.; photosynthetic characteristics; root system characteristics; daily change
2013-09-20
教育部博士点基金“广东石漠化地区优良抗逆树种筛选及植被恢复机理研究”(20124404110007);亚热带农业生物资源保护与利用国家重点实验室开放课题“桉树水分利用效率及影响因子研究”(KSL-CUSAb-2012-07)
邱 权(1989-),男,湖北黄梅人,硕士研究生,主要研究林木栽培生理生态;E-mail:qiuquan89@163.com
李吉跃(1959-),男,四川金坛人,教授,博士,主要研究森林培育及栽培生理生态;E-mail:ljyymy@vip.sina.com
S792.39;S718.43
A
1673-923X(2014)01-0053-07
[本文编校:吴 毅]
猜你喜欢
干旱地区农业研究(2022年1期)2022-01-28
西南农业学报(2021年7期)2021-08-23
节水灌溉(2021年5期)2021-05-26
农业与技术(2020年12期)2020-07-04
中国农业科技导报(2020年12期)2020-03-15
装备制造技术(2019年12期)2019-12-25
农学学报(2019年1期)2019-02-21
科技创新与品牌(2019年12期)2019-02-06
新课程·下旬(2017年7期)2017-08-14
中国果业信息(2015年9期)2015-01-22
