
GHR缺失突变对隐性白羽鸡肌肉组织学特性和 IM F的影响
2013-12-22 06:32:24陈仲灿牟和平郭金彪张绮琼刘为民林树茂
佛山科学技术学院学报(自然科学版) 2013年3期
陈仲灿,牟和平,郭金彪,张绮琼,刘为民,林树茂*
(1.佛山市南海区桂城街道农林服务中心,广东佛山 528251;2.成都天邦生物科技股份有限公司 ,四川 成都 610100;3.佛山科学技术学院 动物科学系 ,广东 佛山 528000)
生长激素受体(Growth Hormone Receptor,GHR)基因的突变是引起性连锁矮小鸡(Sex-linked Dwarf Chicken,SLD)的主要原因,其突变方式包括点突变、结构基因突变、GHR调控区突变以及缺失突变等,而且不同突变类型所导致的性连锁矮小型鸡在生产性能上可能表现出较大的差异[1-5]。通过RFLP分析,发现正常鸡GHR基因的一个6.0 kb条带在矮小鸡仅为 4.3 kb,认为这一出现于外显子 10和 3′UTR区的 1.7 kb缺失是导致矮小鸡的原因[1]。
在优质肉鸡育种中,隐性白羽鸡发挥了重要作用。实际上,该品种是是从白洛克(或白温多得)中选育出来的一种快大白羽肉鸡中的某些品系。在长期的育种实践中,在隐性白羽鸡的基础上,培育了一个携带GHR缺失突变的矮小型隐性白羽鸡。研究表明,GHR基因的缺失突变会引起肌纤维数量减少和肌纤维直径变小,使矮小鸡骨骼及生长发育受阻[6]。
肌内脂肪(intramuscular fat,IM F)是沉积在肌肉块之内的脂肪,分布于肌外膜、肌束膜以及肌内膜上。当肌束和肌纤维间沉积一定脂肪时,构成肉品的大理石纹状,它不仅提高肌肉的感观满意程度,而且增强肉的风味、嫩度以及多汁性[7]。对哺乳动物多能干细胞系的研究表明,脂肪细胞来源于间充质干细胞[8-10],它们经过多次的细胞代谢分化为前脂肪细胞、不成熟脂肪细胞,最终形成成熟脂肪细胞。
为了进一步研究GHR缺失突变对肌肉组织学特性和肌内脂肪(IM F)沉积的影响,以GHR缺失突变导致的生长发育受阻的矮小型隐性白羽鸡为研究对象,以正常型鸡为参照,对其7周龄的肌肉组织学特性和肌内脂肪含量进行了测定,旨在为鸡的育种实践提供理论参考。
1 材料与方法
1.1 实验动物
试验所用的2个品种均为隐性白羽洛克鸡。其中1个以隐性白羽洛克鸡为基础,通过导入矮小型基因(dw)并经多个世代的选育,形成了遗传性能稳定的矮小型隐性白羽洛克鸡,简称矮小型鸡;另 1个为体型正常的隐性白羽洛克鸡,简称正常型鸡。遗传分析表明,矮小型鸡属于GHR基因外显子10末端和3’UT R的1.73 kb大片段缺失突变,未发现编码区的剪切位点突变;正常型鸡则未发现GHR基因存在突变。
1.2 饲养过程与方法
在进小鸡的前一周对鸡舍的地面和墙壁、料槽、饮水器进行全面的清洗、消毒后,再用高锰酸钾和福尔马林对鸡舍进行密闭熏蒸消毒(每 m3空间用 20 g高锰酸钾、40 m L福尔马林),并准备好保温灯,垫料,照明器材等育雏设备。育雏室采用红外线灯泡保温,第1周保持32~ 35°C,次后每周降 2°C,直至脱温。采用室内笼养与自由采食相结合的饲养方法,饲养期为84 d。每天早、中、晚饲喂3次,每次把喂料量记录下来同时也把死亡的个体数及时记录下来。每周四早上进行空腹称重并记录下来。试验所用饲料的营养水平见表1。
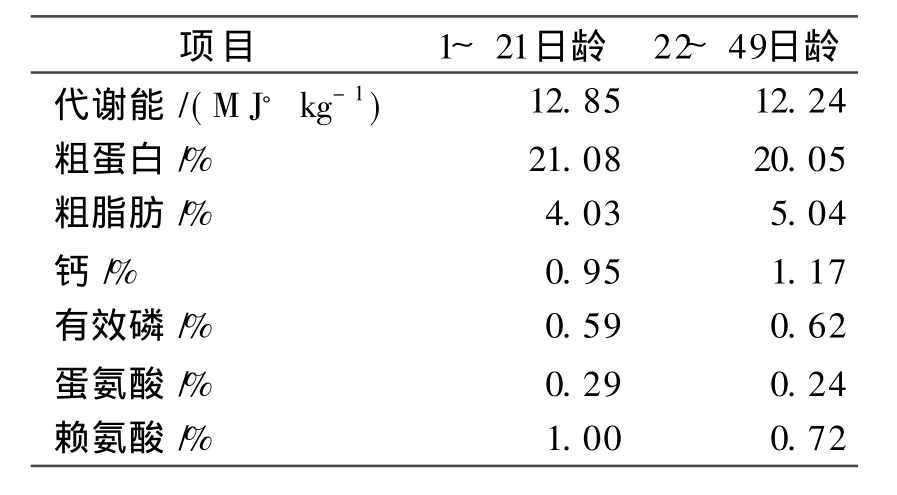
表1 不同阶段肉鸡的营养水平
1.3 样本的采集
随机抽取按常规饲养方法饲养至 7周龄的矮小型鸡和正常型鸡母鸡各9只,分离其腿部肌肉。
1.4 肌内脂肪含量的测定
采用索氏抽提法测定肌内脂肪含量,参考《肉与肉制品游离脂肪含量的测定》(国标 GB/T-6434-2006),使用乙醚抽提单块肌肉待测样脂肪。每个样品测定3次,以平均值作为样品的肌内脂肪含量。根据公式 IM F=(提前总重-提后总重)/提前样品干物质总重×100%,计算干物质中 IM F的含量。
1.5 肌肉组织学特性的测定
试验鸡放血后,立刻取腓肠肌放入冰冻切片机冷冻(-21°C)30 min后切片,切片厚度为 16μm,每个样品做 3张切片,然后将切片进行常规 HE染色。每个样品的 3张切片中,在 10×20倍视野下,随机选取 5个视野,Nikon E4500数码相机拍照,然后通过江苏捷达科技形态学分析软件,分别计数肌纤维横切面的长轴长度、短轴长度、肌纤维根数、肌纤维间距、肌大束间距。再由长轴长、短轴长的几何平均数得到肌纤维直径;由肌纤维根数、视野面积两者相除得到肌纤维密度。
2 结果
2.1 矮小型鸡与正常型鸡IMF含量的测量结果
肌内脂肪 (IMF)含量,矮小型鸡平均 14.28%,正常型鸡 11.85%。矮小型鸡极显著大于正常型鸡(P<0.01),表明矮小型鸡肌内脂肪沉积能力比正常型鸡更强,如表2所示。表2中,数字右上角字母不同为差异极显著(P<0.01),字母相同为差异不显著(P> 0.05),表3同。

表2 两个品种鸡腿肌中肌内脂肪(IM F)含量 %
2.2 矮小型鸡与正常型鸡肌纤维组织学特性的测量结果
经测量,肌纤维直径,正常型鸡平均 59.28μ m,矮小型鸡平均 49.07μ m,矮小型鸡极显著小于正常型鸡(P<0.01);肌纤维密度,正常型鸡平均318根 /mm2,矮小型鸡平均421根 /mm2,矮小型鸡显著高于正常型鸡(P<0.01);肌纤维间距和肌大束间距,正常型鸡平均分别为 5.96μ m和 11.64μ m,矮小型鸡平均分别为 5.78μm和 11.85μ m,两者差异不显著(P> 0.05),如表3所示。
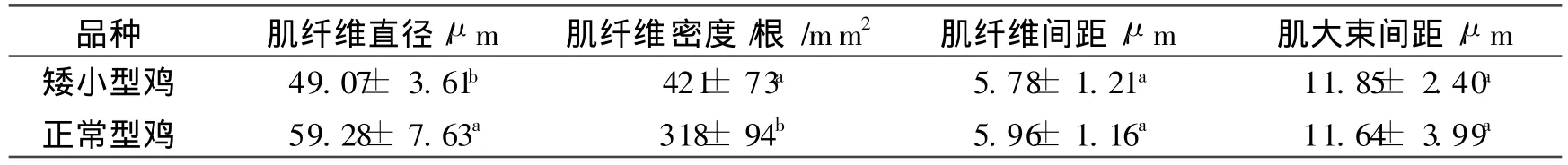
表3 两个品种鸡肌纤维组织学特性
图 1和图 2为两个品种鸡的骨骼肌冰冻切片 HE染色的结果。
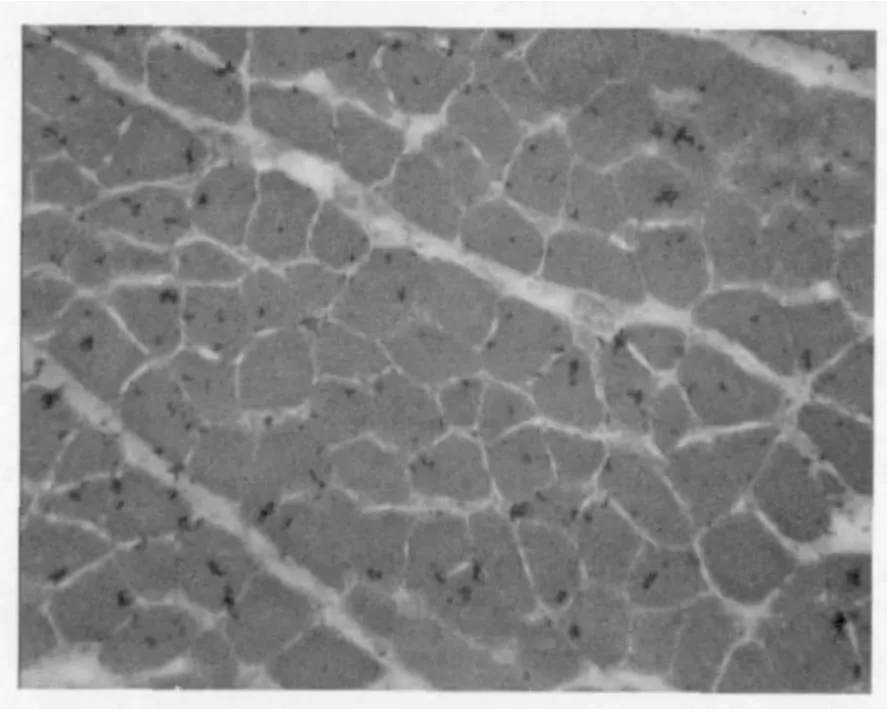
图1 正常型鸡骨骼肌冰冻切片(HE染色 200×)
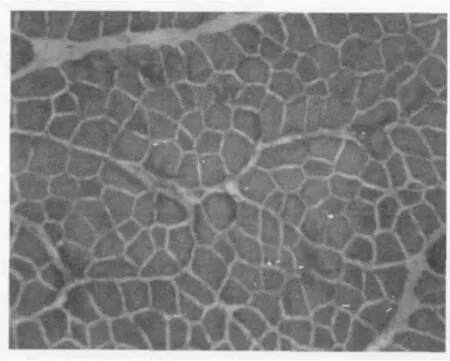
图2 矮小型鸡骨骼肌冰冻切片(HE染色 200×)
3 讨论
3.1 GHR缺失突变使隐性白羽鸡肌内脂肪沉积能力增强
从研究结果来看,GHR缺失突变所引起的矮小型隐性白羽鸡的 IMF含量极显著高于正常型鸡,表明矮小型鸡的肌内脂肪沉积能力增强。关于矮小基因导致动物体内脂肪含量升高,以往有许多类似的报道。早在 1971年,GUILLAUME[11]在对鸡dw基因的作用进行综述时就曾提到矮小鸡比正常鸡具有更高的脂肪含量。 TOUCHBURN[12]等研究了性连锁矮小基因(dw)对5周龄矮小型鸡屠体脂质积累的影响,结果发现矮小型鸡的屠体脂肪含量比非矮小型鸡明显增加,认为脂质沉积量的增加是由于脂肪合成增加和能量消耗减少引起的。BURGHELLE-MAYEUR[13]等证实矮小鸡由于脂肪组织的脂肪分解能力下降,使得脂肪含量增加。 SANGIAO-ALVARELLOS[14]等指出,GH缺乏症(GHD)患者和GHR缺乏的小鼠由于脂肪分配和脂肪动员的变化引起脂肪比例增加。大部分肉用畜禽都具有通过增加脂肪组织,而不是增加蛋白组织来表达增重的潜力。近年来,多数研究结果表明,肉鸡胴体脂肪含量过多是由于对增重和生长速度的选择所致,用小白鼠进行的许多研究,也支持这种结论[15-17]。此外,改善大白鼠和小白鼠饲料转化率的选择会增加胴体脂肪。因此,这些研究的总的结论是体重、生长速度和饲料转化率的遗传改进都会导致胴体脂肪过多[18]。
3.2 GHR缺失突变对隐性白羽鸡肌肉组织学特性和肌肉品质的影响
研究结果显示,GHR缺失突变所引起的矮小型鸡的肌纤维密度显著高于正常型鸡,与之相对应,矮小型鸡肌纤维直径极显著低于正常型鸡。一般情况下,肌纤维密度与肌纤维直径呈负相关,肌纤维直径越小,单位面积所含肌纤维越多,肌纤维密度越大。
另外,影响肌纤维粗细的因素还包括年龄、品种、营养状况和运动量等。年龄越大,肌纤维越粗;地方品种的肌纤维直径较改良品种的肌纤维直径细;营养水平低时,肌纤维直径较小;日增重越快,肌纤维直径增粗较快。
许多研究表明,肌纤维越细,肉质越嫩;肌纤维越粗,肉质越差,肌纤维直径与肉品的柔嫩度密切相关。孙竹珑[19]对藏山羊的组织学特性研究发现,藏山羊肌纤维直径小,肉质细嫩,品质良好;肌纤维越细、密度越大的品种,其肌内脂肪的沉积量要多于肌纤维粗而密度低的品种。也就是说,肌纤维直径越小,密度越大,则肌内脂肪越多,肉质越好。黄涛[20]曾以感官指标与肌肉组织学指标相结合的方法评定鸡肉品质,其中感官指标是以专业食品评价员进行评定,最终结果为肌纤维密度越大,脂肪含量越高,鸡肉品质越好。
研究表明,骨骼肌肌纤维越细,肌束内肌纤维越密,肉质也越嫩。动物出生后肌肉体积的增加主要是由于已经存在的肌纤维体积的增大,随着肌纤维的增粗,肌肉间结缔组织与脂肪组织的增加,肌纤维密度下降。吴信生等[21]对我国7种地方鸡种的组织学研究表明,肌纤维密度与肌肉的嫩度相关比较密切,7个鸡种均表现为肌纤维密度越大,肉质越细嫩,7个地方鸡种肌纤维密度与嫩度呈负相关,肌纤维直径与嫩度呈正相关,即肌纤维愈细,肌纤维密度愈大,肉质越鲜嫩。王亚鸣等[22]研究发现,骨骼肌纤维越细,肌纤维密度越大,肌肉脂肪含量高,则肉质细嫩、味美多汁。矮小型鸡的肌纤维间距、肌大束间距与正常型鸡差异不显著。由于肌肉内的脂肪组织主要分布在肌束和肌纤维之间,肌内脂肪的沉积使肉表现出大理石纹,肌大束间距较大,这有利于沉积脂肪,使肌内脂含量达到理想水平,从而提高肉的风味[6]。
4 小结
GHR基因缺失突变所引起的矮小型隐性白羽鸡,肌内脂肪沉积能力高于正常型鸡,肌纤维直径极显著降低,肌纤维密度极显著升高,表明矮小型隐性白羽鸡的肌肉组织学特性和脂肪沉积能力发生重大变化。GHR缺失突变所引起的上述变化对改善隐性白羽鸡的肌肉品质有利。
[1] AGARWAL S K,COGBURN L A,BURNSIDE J.Dysfunctional growth hormone receptor in a strain of sex-li nked dwarf chicken:evidence for a mutation in the intracellular domain[J].J Endocrinol,1994,142(3):427-434.
[2] HUANG N,COGBURN L A,AGARWAL S K,et al.Overexpression of a truncated growth hormone receptor in the sex-linked dwarf chicken:evidence for a splice mutation[J].M ol Endocrinol,1993,7(11):1391-1398.
[3] TAN AKA M,HAYASHIDA Y,W AKITA M,et al.Expression of aberrantly spliced growth hormone receptor mRN A in the sex-linked dwarf chicken,Gifu 20[J].Growth Regul,1995,5(4):218-223.
[4] ROSEN FELD R G,ROSENBLOOM A L,GUEV ARA-Aguirre J.Growth hormone(GH)insensitivity due to primary GH receptor deficiency[J].Endocr Rev,1994,15(3):369-390.
[5] 肖璐 ,李宁,戴茹娟 ,等.性连锁矮小鸡(dwdw)生长激素受体(cGHR)基因突变的精确定位 [J].农业生物技术学报,1996,4(2):174-178.
[7] NO RBERT H,HAUN ERLAND S,FRIEDRICH S.Fatty acid-binding proteins-insights from genetic manipulations[J].Prog Lipid Res,2004,43(4):328-349.
[8] TON G Q,HOT AMISLIGIL G S.Molecular mechanisms of adipocyte differentiation[J].Rev Endocr Metab Disord,2001,2(4):349-355.
[9] ROSEN E D,M ACDOUGALD O A.Adipocyte differentiation from the inside out[J].Nat Rev Mol Cell Biol,2006,7(12):885-896.
[10] FARMER S R.Transcriptional control of adipocyte formation[J].Cell Metab,2006,4(4):263-273.
[11] GUILLAUME J.Some nutritional and physiological traits of the dw chick[J].Worlds Poult Sci J,1971,27(3):284-285.
[12] TOUCHBURN S P,GUILLAUME J,LECLERCQ B,et al.Lipid and energy metabolism in chicks affected by dwarfism(dw)and Naked-neck(Na)[J].Poult Sci,1980,59(10):2189-2197.
[13] BURGHELLE-M AYEU R C,TIXIER-BOICHARD M,M ERAT P,et al.De novo lipogenesis and lipolysis activities in normal(Dw)and dwarf(dw)White Leghorn laying hens[J].Comp Biochem Physiol B,1989,93(4):773-779.
[14] SANGIAO-ALV ARELLOS S,VZQUEZ M J,VARELA L,NOGUEIRAS R,et al.Central ghrelin regulates peripheral lipid metabolism in a growth hormone-independent fashion[J].Endocrinology,2009,150(10):4562-4574.
[15] 陈继兰,文杰 ,赵桂苹,等.鸡肉肌苷酸和肌内脂肪等肉品风味性状遗传参数的估计[J].遗传,2005,27(6):898-902.
[16] 黄秀清,黄建斌,翁志铿,等.5种不同类型地方鸡种肉用性能及肉质的研究 [J].福建农业学报 ,2000(1):35-39.
[17] 李文娟,李宏宾,文杰,等.鸡 H-FABP和 A-FABP基因表达与肌内脂肪含量相关研究 [J].畜牧兽医学报,2006,5(7):417-423.
[18] 张海艳,于太永,关伟军.肌苷酸形成机理及其含量影响因素浅析 [J].中国农业科技导报,2004,6(3):17-21.
[19] 孙竹珑.藏山羊肌肉组织学特性研究[J].西南民族学院学报:自然科学版,1991,17(3):48-54.
[20] 黄涛,刘华贵,徐淑芳,等.相同营养条件下五种鸡肉肌苷酸及其相关代谢物含量的比较研究[J].饲料工业,2005,26(10):45-47.
[21] 吴信生,陈国宏,陈宽维,等.中国部分地方鸡种肌肉组织学特点及其肉品质的比较研究 [J].江苏农学院学报,1998,19(4):52-58.
[22] 王亚鸣,刘龙芳.江西玉山猪肌肉组织学特征与肉质的关系 [J].江西农业大学学报,1994,16(3):284-287.
猜你喜欢
科普童话·学霸日记(2023年6期)2023-08-21 09:07:08
中国临床医学影像杂志(2022年6期)2022-07-26 07:17:24
中国临床医学影像杂志(2022年5期)2022-07-26 07:11:54
肝博士(2021年1期)2021-03-29 02:32:08
国际放射医学核医学杂志(2021年10期)2021-02-28 08:43:54
动漫界·幼教365(小班)(2019年3期)2019-06-11 05:27:57
Communications in Theoretical Physics(2017年6期)2017-05-12 08:53:09
天津农学院学报(2016年2期)2016-12-01 05:40:05
成都中医药大学学报(教育科学版)(2016年1期)2016-01-22 07:24:50
实用肝脏病杂志(2015年5期)2015-12-03 06:28:04
