
香鱼RBP基因的克隆、组织表达特征及其与盐度应激相关的表达分析
2013-12-21 07:52史雨红
生物学杂志 2013年3期
陈 强, 赵 桐, 史雨红, 陈 炯
(宁波大学应用海洋生物技术教育部重点实验室,浙江 宁波315211)
视黄醇结合蛋白(Retinol binding protein,RBP)最早分离自人类血浆,是脂质转运蛋白(lipocalin)家族成员,为一种低分子量的载体蛋白[1]。RBP存在血浆和细胞内两种形式。血浆RBP主要是将全反式视黄醇(即维生素A,Vitamin A)及其衍生物从肝脏转运至靶组织;细胞内RBP参与细胞内视黄醇的转运和代谢[2]。近来,研究发现RBP与脂类代谢相关,尤其是与肝酯酶(hepatic lipase,HL) 活性紧密相关[3],而且血浆中RBP和甲状腺素蛋白(transthyretin,TTR)的蛋白比例可作为体内脂类代谢的生化指标[4]。最新研究表明,RBP-视黄醇复合物通过与脂肪细胞表面受体视磺酸激活基因6(stimulated by retinoic acid 6,STRA6)结合,可激活细胞内与脂类积累相关基因表达,如过氧化物酶体增殖物活化受体γ (peroxisome proliferator-activated receptor γ,PPARγ)[5]。与哺乳动物相比,鱼类RBP研究较少,目前已有斑马鱼(Daniorerio)[6]、金头鲷(Sparusaurata)[7]、鲤鱼(Cyprinuscarpio)[8]等多种鱼类RBP基因序列被测定。据已有文献报道,鱼类RBP的功能主要与繁殖、胚胎发育相关[9-11]。
香鱼主要分布于中国渤海、黄海、东海及台湾省的入海溪流中,为一年生广盐性洄游鱼类。与其它鱼类相同,香鱼主要由肾、鳃等组织进行渗透调节,维持组织内特定的水分和盐类平衡,以适应不同盐度的环境,因此香鱼可作为研究鱼类渗透压调节机制和生理学基础理论的模型。鱼类盐度应激反应机制的研究,对提高鱼类渗透压调节能力,降低海水驯养或淡水驯化鱼类的生产成本,以及在养殖过程中控制适宜的水体盐度与采用合理的盐度调节模式具有重要的经济意义和科学指导意义。
我们前期双向电泳实验揭示,香鱼在盐度适应过程中脂类代谢相关基因表达显著变化,其中肾脏RBP表达量下降[12]。为了进一步解析RBP在香鱼盐度应激适应中的作用,本文克隆了香鱼RBP基因cDNA全序列,明确了其mRNA的组织表达特征,并分析了盐度变化过程中RBP基因mRNA和血浆中蛋白表达的变化。
1 材料与方法
1.1 实验材料和试剂
引物由上海英俊生物有限公司合成;总RNA抽提试剂RNAiso、文库构建相关试剂盒、RNAiso试剂和SYBR Premix Ex Taq试剂盒均购自Takara公司;Gel Extraction Kit 试剂盒购自Omega公司;硝酸纤维素膜(0.45μm)购自millipore公司;二抗(辣根酶标记山羊抗小鼠IgG)购自北京中山金桥生物技术有限公司;ECL发光试剂盒、显影定影试剂盒、柯达X-OMAT BT胶卷购自碧云天生物技术研究所。
1.2 盐度应激实验
实验所用香鱼体重80~100 g、体长12~15 cm,购自宁波水产大世界。实验期间定期投喂饲料,不间断充气,控制水温为20℃±1℃。实验设置4组:盐度0、盐度5、盐度10和盐度15,每组4条鱼。在淡水中暂养2个星期后,咸淡水组采用3 d内逐步升高盐度的方式到达指定盐度。3个星期后分别取两组香鱼的肝、脾、肾、肠、脑、心、鳃、肌肉共8个组织样品,现场液氮速冻,-80℃冰箱保存。不同处理组香鱼尾静脉抽血后于4 ℃静置4 h,冷冻离心取上清备用。
1.3 cDNA序列获得及分析
根据已报道的鱼类RBP保守序列设计简并引物dRBPf(+): 5′-TGGACRG HTAYTCCTTCRTCTT-3′ (R=A,G;H=A,T,C;Y=T,C)。采用RACE技术从已构建的香鱼肝脏cDNA文库中获得RBP的cDNA序列。序列拼接后用BLASTX (http://www.ncbi.nlm. nih.gov/BLAST/)分析。信号肽序列预测采用SignalP 4.0程序[13]。系统进化树分析采用MEGA 4.0软件[14]。
1.4 实时荧光定量RT-PCR (RT-qPCR)法检测组织表达特征
根据香鱼RBP全长基因序列设计特异引物aRBP(+):5′-GATGACAGCCTCTGCCCAG-3′和aRBP(-):5′-CACAGAAACCTGTGTGTGGG-3′,预期扩增片段长368 bp;内参采用香鱼β-actin看家基因,扩增引物为pActin2 (+)5′-TCGTGCGTGACATCAAGGAG-3′和pActin2 (-): 5′-CGCACTTCATGATGCTGTTG-3′,预期扩增片段长231 bp。采用RNAiso试剂提取健康香鱼各组织总RNA,DNase I处理、第一链cDNA合成和PCR扩增等方法同文献[15]。荧光定量的结果用SPSS软件(15.0)中的单因素方差分析法(ANOVA)进行分析。
1.5 RT-qPCR检测盐度升高时RBP基因mRNA表达变化
不同盐度应激的组织样本总RNA抽提、第一链cDNA合成,RT-qPCR的引物序列同1.4。盐度0、盐度5、盐度10和盐度15各组生物学重复n=4,技术重复3次。
1.6 原核表达、抗血清制备和Western blot分析
根据RBP基因cDNA序列设计原核表达引物pET-aRBP(+)(5′-GGAATTCGACTGTCAG GTTGCCAACAT-3′)和pET-aRBP(-)(5′-CCTCGAGTTAACTGCTCTCACAGAAAC-3′)(下划线表示限制性内切酶EcoR I和XhoI酶切位点)。重组质粒pET28a-aRBP的构建,诱导表达、抗血清制备及Western blot方法同文献[15]。
2 结果
2.1 香鱼RBP基因cDNA序列分析
采用两次RACE后序列拼接获得了包含有完整阅读框的香鱼RBP序列(FN392685)。核苷酸全序列分析表明,香鱼RBP基因cDNA序列长为932 bp(不计polyA尾)。开放阅读框共576 bp (58nts-633nts,含终止密码子),编码一个由192个氨基酸组成的蛋白,其分子量约21.9 kDa,其等电点pI为6.08。前16个氨基酸残基为信号肽序列。将香鱼RBP氨基酸序列与其它已知的RBP氨基酸序列进行多序列比对分析发现,RBP一级结构中存在有6个保守的半胱氨酸残基,组成3对二硫键,在空间上形成结合——分子全反式视黄醇的位点[16]。序列分析表明,动物RBP高度保守,香鱼RBP与胡瓜鱼(Osmerusmordax)的同源性最高,为92.7%,与大西洋鲑(Salmosalar)次之,为85.4%。
基于RBP氨基酸序列的系统进化树分析表明,鱼类RBP在进化上紧密相关,形成一个显著群体;两栖类、鸟类、哺乳类动物编码的RBP在进化上也紧密相关,并且物种亲缘关系高的,在系统进化树中也高度相关(图1)。

图1 基于RBP基因氨基酸序列的NJ系统发生树
分叉处数值表示1 000次重复抽样所得到的置信度百分比,只显示置信度60 %以上的数值; 标尺长度表明每个位点发生0.1次置换。所用序列登录号为:香鱼(FN392685),胡瓜鱼(BT075450),大西洋鲑(BT048297),虹鳟(AF538329),斑马鱼(NM_130920),金头鲷(AF083556),白斑狗鱼(BT079072),鲤鱼 (AJ277123),黑青斑东方鲀(CAAE01015106),鸡(NM_205238),马苏麻哈鱼(EU325861),人(AL356214),牛(BC109815),猪(NM_214057),小鼠(NM_011255),非洲爪蟾(J02718),欧洲海鲈(FQ310507),蓝鲶(GU588259),尼罗罗非鱼(XM_003441859),褐石斑鱼(JF501991),斜带石斑鱼(FJ438490)。
2.2 香鱼RBP基因 mRNA的组织表达特征分析
RT-qPCR分析结果显示,RBP基因mRNA在肝脏中表达量最高,肾、肠、脑和鳃表达次之,肌肉中表达量相对较少,而脾和心中几乎不表达(图2)。
2.3 香鱼RBP基因在盐度升高时mRNA表达量变化
选取对照香鱼中表达RBP基因mRNA的肝、肾、脑、鳃和肠组织进行RT-qPCR检测。RT-qPCR结果表明,5个组织中RBP基因mRNA表达量呈不同程度下降趋势(图3)。脑组织的表达量无显著性变化;肝和肠组织中表达量在盐度15时,有显著下降(P<0.05),分别下降至盐度0时的0.59倍和0.48倍;肾和鳃组织中盐度下降表达量变化最显著(P<0.01),肾组织中盐度升高时,表达量分别下降至盐度0时的0.51倍、0.08倍和0.05倍,鳃组织中表达分别下降为盐度为0时的0.53倍、0.24倍和0.06倍。

图2 RT-qPCR分析RBP mRNA在香鱼组织中的表达情况
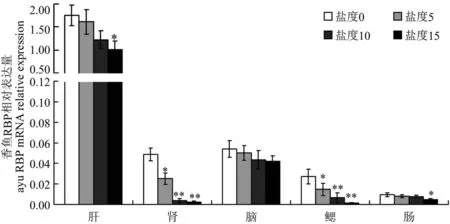
图3 盐度升高时香鱼肝、肾、脑、鳃、肠中RBP 基因mRNA表达量变化

图4 香鱼RBP基因的原核表达(a)和Western blot检测(b)
2.4 香鱼RBP基因的原核表达和抗血清制备
测序验证正确的表达载体pET28a-aRBP在大肠杆菌BL21 Plys E中经IPTG诱导表达,SDS-PAGE电泳分离得到一条表达量很高的蛋白条带,相对分子质量约为24.1 kDa,与预计大小(RBP成熟肽20.3 kDa+融合蛋白3.8 kDa)相近 (图4a)。该片段切胶纯化后免疫小鼠,制备抗血清。用Western blot对获得的抗血清进行验证,结果抗血清具有很高特异性(图4b),可以用于香鱼RBP蛋白含量变化检测。
2.5 Western blot检测香鱼RBP在血清中含量的变化
取不同盐度处理组香鱼血清进行Western blot分析。结果表明,随盐度升高,香鱼血清RBP表达量逐渐下降,盐度5时为盐度0时的0.71倍,盐度10时为盐度0时的0.57倍(P< 0.05) ,盐度15时是盐度0时的0.26倍(P< 0.01) (图5)。

图5 盐度升高时香鱼血清RBP 表达量变化
3 讨论
在本研究中,我们克隆并测定了香鱼RBP基因的cDNA序列,其长度为932bp,开放阅读框由192个氨基酸组成,N端具有信号肽。香鱼RBP与其他已报道的鱼类RBP高度相似,不同物种RBP基因的亲缘关系与物种进化吻合。RBP在其它组织中少量表达,但主要还是在肝脏中表达并释放到血清中而进入各种组织,这与已发表文献报道一致[8]。
当环境盐度改变时,机体需要消耗大量的能量来调节体内渗透压和离子平衡[17]。文献报道,小锯盖鱼(Centropomusparallelus,fat snook)在盐度30时,主要以脂类物质代谢为主要能量代谢形式,以适应高盐环境[18]。香鱼在盐度升高时肾脏中脂类代谢基因表达显著变化[12]。由此说明鱼类在适应盐度变化过程中,脂类代谢加强以提供离子主动运输所需的能量。RBP参与脂类代谢,一方面是由于视黄醇类物质参与调节脂类代谢,包括肝脏和肠甘油三酯的合成和分泌、脂肪酸beta-氧化和脂蛋白C-III合成等[19]。另一方面RBP被认为是新型脂肪细胞因子(adipokine),与脂类物质代谢直接相关。RBP-视黄醇复合物可作为信号分子作用于脂肪细胞表面受体STRA6,导致胞内PPARγ基因的表达[5],而PPARγ基因能调节脂类代谢靶基因的表达,降低血脂水平,限制甘油三酯分解,促进成熟脂肪细胞中脂肪酸的吸收和储存,加速脂肪沉积[20,21]。本研究表明,盐度升高时,肝脏、肾、鳃和肠RBP基因mRNA表达量呈不同程度下降趋势,并且血浆中RBP蛋白也显著下降。我们推测由于RBP的减少,RBP-视黄醇复合物减少,导致STRA6介导的信号通路失效,PPARγ基因的表达受阻,有利于血脂水平的升高和脂肪动员,提高机体能量代谢水平。
综上所述,本研究报道了香鱼RBP基因的克隆及序列分析,并探讨了其表达与环境盐度应激的相关性,为后续蛋白功能的研究奠定了基础,并有助于了解洄游性鱼类盐度适应的分子机理。
[1]Kanai M,Raz A,Goodman D S. Retinol-binding protein: the transport protein for vitamin A in human plasma [J]. J Clin Invest,1968,47(9): 2025-2044.
[2]Bellovino D,Apreda M,Gragnoli S,et al. Vitamin A transport: in vitro models for the study of RBP secretion [J]. Mol Aspects Med,2003,24(6): 411-420.
[3]von Eynatten M,Lepper P M,Liu D,et al. Retinol-binding protein 4 is associated with components of the metabolic syndrome,but not with insulin resistance,in men with type 2 diabetes or coronary artery disease [J]. Diabetologia,2007,50(9): 1930-1937.
[4]Henze A,Frey S K,Raila J,et al. Alterations of retinol-binding protein 4 species in patients with different stages of chronic kidney disease and their relation to lipid parameters [J]. Biochem Biophys Res Commun,2010,393(1): 79-83.
[5]Berry D C,Noy N. Signaling by vitamin A and retinol-binding protein in regulation of insulin responses and lipid homeostasis [J]. Biochim Biophys Acta,2012,1821(1): 168-176.
[6]Sammar M,Babin P J,Durliat M,et al. Retinol binding protein in rainbow trout: molecular properties and mRNA expression in tissues [J]. Gen Comp Endocrinol,2001,123(1): 51-61.
[7]Funkenstein B. Developmental expression,tissue distribution and hormonal regulation of fish (Sparusaurata) serum retinol-binding protein [J]. Comp Biochem Physiol Part B: Biochem Mol Biol,2001,129(2-3): 613-622.
[8]Bellovino D,Morimoto T,Apreda M,et al. Isolation,expression and characterization of carp retinol-binding protein [J]. Gene,2002,295(2): 231-240.
[9]Sammar M,Levi L,Hurvitz A,et al. Studies on retinol-binding protein during vitellogenesis in the Rainbow Trout (Oncorhynchusmykiss) [J]. Gen Comp Endocrinol,2005,141(2): 141-151.
[10]Levi L,Levavi-Sivan B,Lubzens E. Expression of genes associated with retinoid metabolism in the trout ovarian follicle[J]. Biol Reprod,2008,79(3): 570-577.
[11]Belliveau D J,Venkatachalam A B,Thisse C,et al. The duplicated retinol-binding protein 7 (rbp7) genes are differentially transcribed in embryos and adult zebrafish (Daniorerio) [J]. Gene Expr Patterns,2010,10(4-5): 167-176.
[12]Chen J,Wu H Q,Shi Y H,et al. The effects of environmental salinity on trunk kidney proteome of juvenile ayu (Plecoglossusaltivelis) [J]. Comp Biochem Physiol Part D: Genomics Proteomics,2009,4(4): 263-267.
[13]Petersen T N,Brunak S,von Heijne G,et al. Signal P 4.0: discriminating signal peptides from transmembrane regions[J]. Nat Methods,2011,8(10): 785-786.
[14]Tamura K,Dudley J,Nei M,et al. MEGA4: molecular evolutionary genetics analysis (MEGA) software version 4.0 [J]. Mol Biol Evol,2007,24(8): 1596-1599.
[15]黄左安,陈 炯,陆新江,等. 香鱼凝血因子X 基因表达与鳗利斯顿氏菌感染的相关性 [J]. 动物学研究,2011,32(5): 492-498.
[16]Colantuoni V,Romano V,Bensi G,et al. Cloning and sequencing of a full length cDNA coding for human retinol-binding protein [ J ]. Nucleic Acids Res,1983,11 (22): 7769-7776.
[17]Tseng Y C,Hwang P P. Some insights into energy metabolism for osmoregulation in fish [J]. Comp Biochem Physiol C Toxicol Pharmacol,2008,148(4): 419-429.
[18]Rocha A J,Gomes V,Ngan P V,et al. Effects of anionic surfactant and salinity on the bioenergetics of juveniles ofCentropomusparallelus(Poey) [J]. Ecotoxicol Environ Saf,2007,68(3),397-404.
[19]Staels B. Regulation of lipid and lipoprotein metabolism by retinoids [ J ]. J Am Acad Dermatol,2001,45(5): S158-S167.
[20]Schadinger S E,Bucher N L,Schreiber B M,et al. PPARγ2 regulates lipogenesis and lipid accumulation in steatotic hepatocytes [J]. Am J Physiol Endocrinol Metab,2005,288(6): 1195-1205.
[21]Xu C,Wang L L,Liu H Y,et al. A novel dual peroxisome proliferator-activated receptors and agonist with beneficial effects on insulin resistance and lipid metabolism [J]. Biotechnol Lett,2006,28(12): 863-868.
猜你喜欢
现代临床医学(2023年1期)2023-03-24
广东茶业(2019年1期)2019-04-28
中国卫生标准管理(2018年10期)2018-01-29
现代检验医学杂志(2016年5期)2016-08-20
东方考古(2016年0期)2016-07-31
广东海洋大学学报(2015年3期)2015-12-22
华南农业大学学报(2015年5期)2015-12-04
饲料博览(2015年5期)2015-04-04
中国洗涤用品工业(2015年5期)2015-02-28
中国医药科学(2015年15期)2015-02-27
