
不同实验生态环境对海刺猬遮蔽行为的影响
2013-12-16 08:18:46常亚青李云霞罗世滨
生态学报 2013年9期
常亚青,李云霞,罗世滨,赵 冲
(大连海洋大学农业部北方海水增养殖学重点实验室,大连 116023)
动物在漫长的进化过程中,通常会形成多样的行为来适应不断变化的外界环境,以延续物种。海胆的遮蔽行为通常是指海胆利用它们的棘和管足将它们周围环境中的小碎片,如贝壳、水草、大型藻类、石块等吸附到它们的反口面,以躲避捕食者的捕食、避免光照和水中悬浮颗粒对身体的伤害以及降低水流对身体的冲击等,以此来进一步提高对周围环境的适应能力[1-5]。目前的研究认为,该行为的影响因素主要分为自身因素和外界环境因素两大类。不同规格、不同种类的海胆及不同运动状态下的海胆其遮蔽行为存在明显的不同[1,4-8]。而捕食者、光照、紫外线、温度、水流波动、水质、遮蔽物材料等被广泛认为是影响海胆遮蔽行为的主要因素[1,3,9-14]。虽然人类在100多年前就已经观察到海胆的遮蔽行为[2-3],但对于该行为的形成机理和影响机制目前仍然不十分清楚。这与国内外相关研究主要集中于短时间内的环境因子变化刺激对海胆遮蔽行为的影响,而对于较长时间的环境变化(特别是遮蔽环境的变化)对海胆遮蔽行为特征的影响缺乏研究有很大的关系。
海刺猬(Glyptocidaris crenularis),属于棘皮动物门、游在亚门、海胆纲、正形目、疣海胆科,是疣海胆科目前仅存的唯一代表种,主要分布在中国黄海北部及日本海的部分海域[15]。先前研究证明,海刺猬具有明显的遮蔽行为,是研究该行为的非常好的实验材料[16]。在实验室条件下,研究了(较长时间)不同遮蔽环境条件对海刺猬遮蔽行为特征的影响,以期进一步深入了解该行为的形成机制、特点和影响因素。
1 材料方法
1.1 实验动物
海刺猬(G.crenularis)于2011年2月购自辽宁大连海宝渔业有限公司,该批海刺猬育于2010年5月。实验前,将海刺猬在实验室300 L水槽中暂养,投喂海带。
1.2 实验设计
饲养实验于2011年3月2日开始,在实验室内进行。实验分为3组:掩蔽组(以累积的空心砖作为掩蔽环境,空心砖的正方形孔径边长为(38.30±3.34)mm,层高为(171.90±7.02)mm、遮蔽组(以菲律宾蛤仔Ruditapes philippinarum的贝壳作为遮蔽环境)和空白组(没有遮蔽物和掩蔽物的环境),每个组4个重复,每个重复20个海刺猬,分别于300 L的水槽中饲养,每个组海刺猬的初始大小规格一样,每个水槽的位置随机分配。实验过程中用海带投喂海刺猬,投喂过程中,砖块组则在砖块内外都放置海带,以保证海胆不因摄食而被迫离开自身的偏好环境。水槽内的海水用气泵正常供应空气,且每3d换1次海水。
到2011年10月6日,即海刺猬在各自的环境中饲养了7个月后,开始进行遮蔽行为观察。实验在室内弱光条件下进行,从每个组的80个海刺猬中分别挑出大小规格类似的海刺猬36个,3个组共108个。每个组分3个重复,每个重复12个海刺猬,分别放置在60 L的水槽中,每个水槽中事先放入一定规格的贻贝(Mytilus edulis)和菲律宾蛤仔(R.philippinarum)的贝壳。整个观察过程持续3 h,每10 min观察1次。观察的内容包括:①每个重复前3个海刺猬分别初次遮蔽自己的耗时,此项为实验开始时观察;② 每个重复有遮蔽行为的海刺猬总数;③每个重复中被海刺猬用于遮蔽的贝壳总数;④每个重复中分别只以贻贝贝壳或者菲律宾蛤仔贝壳作为遮蔽物的海刺猬数,此项每1 h观察1次。
1.3 统计方法
利用Excel和SPSS统计分析软件对数据进行整理和分析,各数据均符合正态性和方差齐性。利用单因素方差分析(One-way ANOVA)统计了观察前各组海刺猬和贝壳规格大小,并比较了各组海刺猬规格和贝壳规格之间的差异性,以确定它们之间没有显著性的差异。利用单因素方差分析比较了各组海刺猬前3个海胆分别初次遮蔽耗时之间的差异。用重复度量分析方法比较了各组海刺猬达到平稳状态后被海刺猬用于遮蔽的贝壳总数和有遮蔽行为的海刺猬总数之间的差异。利用双因素重复度量方差分析比较了3种遮蔽环境下海刺猬分别只以贻贝贝壳或者菲律宾蛤仔贝壳作为遮蔽物的海刺猬数量的差别,发现环境因素和贝壳选择之间的交互作用并不显著。因此,进一步比较了在不考虑环境因素情况下海刺猬分别只以贻贝贝壳或者菲律宾蛤仔贝壳作为遮蔽物的海刺猬数量的差异性。
2 结果
实验各组所用贝壳的壳长、壳高和壳重的均值及标准差列于表1,两种贝壳的规格差异较显著。实验所用各组海刺猬的壳径大小没有显著性的差异(P>0.05)(表1)。
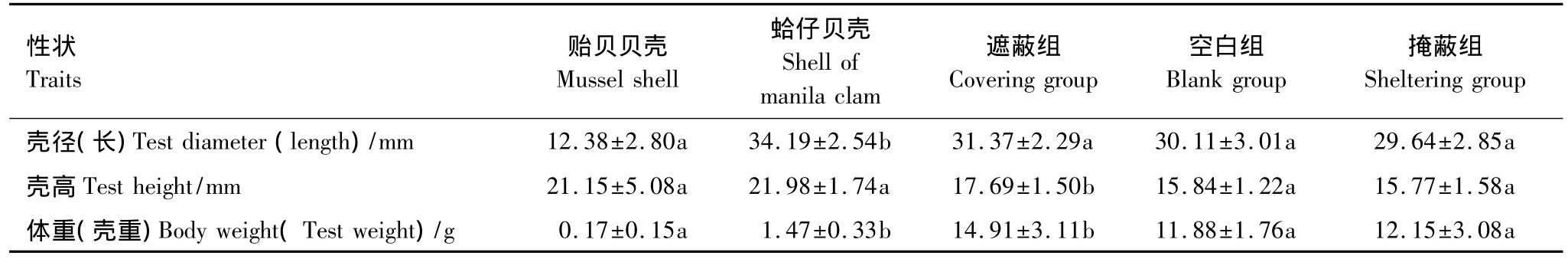
表1 遮蔽材料及各组海刺猬的体尺性状Table 1 Covering materials and body size traits of sea urchins(G.crenularis)
单因素方差分析(One-way ANOVA)表明,遮蔽组和掩蔽组,前3个海刺猬分别初次遮蔽自己的耗时要显著短于空白组(P<0.05),而遮蔽组和掩蔽组之间没有显著性的差异(P>0.05)(表2)。

表2 海刺猬初次遮蔽时长的方差分析Table 2 The One-way ANOVA of the time when sea urchins(G.crenularis)first covered themselves
3组海刺猬,其有遮蔽行为的海胆总数随着时间的变化,总体都呈现先增加后趋于平稳的趋势,但是3个组有遮蔽行为的海刺猬总数到达稳定状态的时间以及平稳后有遮蔽行为的海刺猬总数有所不同(图1、图2)。遮蔽组的海刺猬在40 min左右达到平稳状态,而掩蔽组和空白组的海刺猬则分别在160 min和40 min左右达到平稳状态。各组都到达稳定状态后,掩蔽组有遮蔽行为的海刺猬总数要显著多于空白组。而遮蔽组稳定后有遮蔽行为的海刺猬总数,要少于掩蔽组,而多于空白组,但是差异不显著(P>0.05)。
3个组被海刺猬用于遮蔽的贝壳总数随时间的变化规律和各组有遮蔽行为的海胆总数的变化规律相似,都呈现先增加后趋于平稳的状态(图1、图2)。遮蔽组的海刺猬在40 min左右达到平稳状态,而掩蔽组和空白组的海刺猬则分别在120 min和40 min左右达到平稳状态。各组都到达稳定状态后,掩蔽组被海胆用于遮蔽的贝壳总数多于遮蔽组和空白组,而遮蔽组稳定后被海刺猬用于遮蔽的贝壳总数要多于空白组,但是它们差异不显著(P>0.05)。
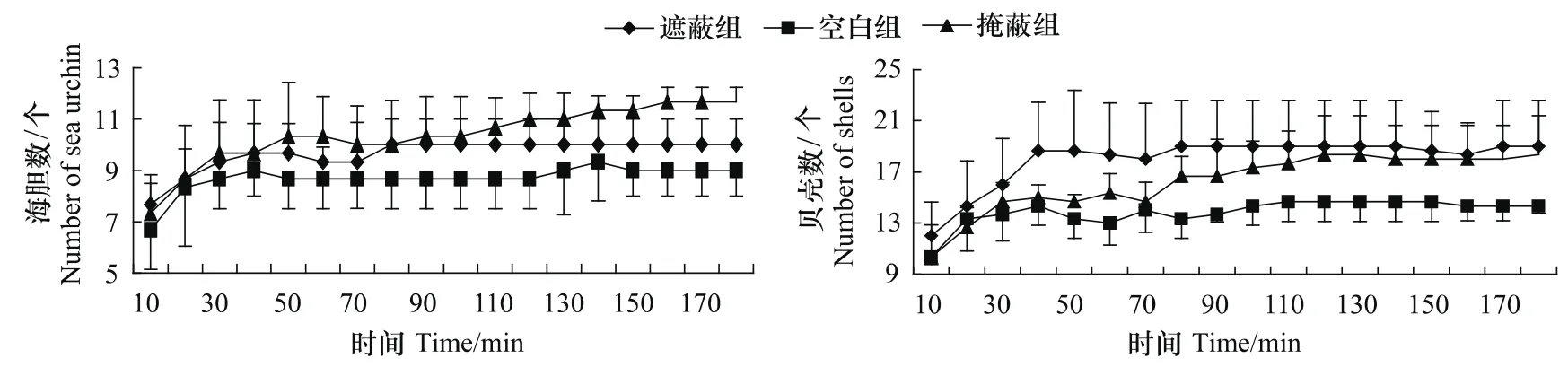
图1 遮蔽的海刺猬总数和被用于遮蔽的贝壳总数随时间的变化规律Fig.1 Number of sea urchins(G.crenularis)covered themselves and total number of shells used for covering during the period of the experiment
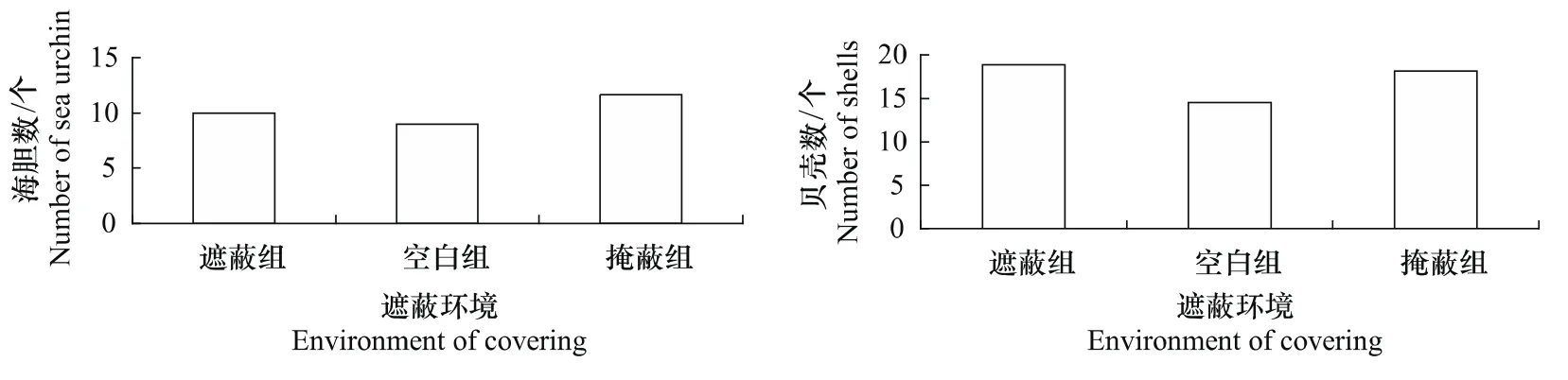
图2 平稳后各组遮蔽的海刺猬总数和被用于遮蔽的贝壳总数Fig.2 Number of sea urchins(G.crenularis)covered themselves and number of shells used for covering
双因素方差分析显示,海刺猬对其遮蔽所利用的贝壳种类具有选择性(P<0.05),而不同的遮蔽条件下的生活环境对海刺猬的这种选择性没有影响(P>0.05)(表3)。因此,去除生活环境因素后,进一步方差分析显示,单独以菲律宾蛤的仔贝壳作为遮蔽物的海刺猬数量要显著多于单独以贻贝的贝壳作为遮蔽物的海刺猬数量(表4)。单独以贻贝贝壳或者菲律宾蛤仔贝壳作为遮蔽物的海刺猬数量要显著的多于同时以两种贝壳作为遮蔽物的海刺猬数量,占到了78.57%。

表3 以一种贝壳作为遮蔽物的海刺猬数量的双因素方差分析Table 3 Two-way analysis of variance of the number of sea urchins(G.crenularis)only used one type shell as covering material

表4 以一种贝壳作为遮蔽物的海刺猬数量的重复度量方差分析Table 4 Repeated measure analysis of the number of sea urchins(G.crenularis)only used one type shell as covering material
3 讨论
有研究发现海胆对光照非常敏感[11],尤其是紫外线,作为海胆遮蔽行为的一个影响因素已经被广泛的研究,被认为是引起数种海胆产生遮蔽行为的主要因素[3,9-10,12]。光照,特别是紫外线,能够破坏动物身体的组织和细胞结构,对动物自身造成伤害。大量研究认为,海胆遮蔽自己的一个主要目的就是为了防止光照对它造成伤害[3,9,13-14,17]。在海刺猬的饲养过程中,掩蔽组的海刺猬很少活动到砖块之外有光照的地方,该组有遮蔽行为的海刺猬总数要显著多于受长期光照影响的空白组,被海刺猬用于遮蔽的贝壳总数也要稍多于空白组。这说明光照能够影响海刺猬的分布,而且长期生活在黑暗条件下的海刺猬再次暴露在光照条件下受光照的影响要大。长期生活在有光照条件下的空白组海刺猬,其遮蔽强度相对于长期生活在无光照条件下的掩蔽组的海刺猬要低很多,而且初次遮蔽自己的速度也慢很多。这说明生活环境中长期光照会使海刺猬的遮蔽行为减弱,这与上述提到的众多研究结论不同,这些研究认为光照能够增强海胆的遮蔽行为的强度。这种不同主要是由于研究方法的不同引起的,先前的研究主要是专注于不同的光照改变强度对海胆遮蔽行为的影响,在光照刺激之前,各处理组的海胆生活环境中的光照条件及状态都是一样的,而观察时的光照条件不一样。本实验主要是专注于生活环境中长期的光照改变对海刺猬遮蔽行为的影响,各处理组的海胆进行遮蔽行为观察之前的生活环境中的光照条件不同,而观察时的光照条件一样。通过与前人实验结论的对比,推测在不断变化着的光照环境当中,海胆的遮蔽行为要更显著,且光照环境的改变量越大遮蔽行为的改变就越显著,这种行为似乎更多的是为了适应一个动态变化的光照环境,而一旦长期处于较稳定的光照环境当中,海胆的遮蔽行为就会减弱。生物对生活的环境会逐渐的产生适应性,这也是生物不断进化的原因之一。在稳定光照环境下海刺猬遮蔽行为变弱的一个很大原因可能就是长期生活在遮蔽及掩蔽物缺失和光照环境下的海刺猬通过提高对光照的耐受力来应对遮蔽及掩蔽条件的缺失,从而对该环境产生了一定的适应性。这表明在海刺猬长期的进化过程中,可以形成多种策略来应对光照对它们的不利影响。但是,掩蔽组的海刺猬,其被海胆用于遮蔽的贝壳总数和有遮蔽行为的海胆总数到达稳定状态的时间要远远长于空白组,这种现象和Verling[3]在拟球海胆(Paracentrotus lividus)中观察到的相类似,这似乎又说明了在黑暗条件生活的海刺猬对光的敏感度下降了。目前还无法解释这一现象,一个可能的原因是长期生活在砖块内黑暗环境下的海刺猬的运动能力减弱了,从而影响了海刺猬初次遮蔽自己的整体速度。
动物的大部分行为都是为了应对外界环境的变化而长期形成的比较稳定的可遗传的行为[1]。海胆的遮蔽行为也被认为是一种长期进化形成的,它有很多作用,既可以伪装自己免受捕食者的伤害[18],也可以减弱阳光对其造成的伤害[3,9,19],增加自己的存活概率。长期生活在遮蔽组和空白组的海刺猬,它们之间有遮蔽行为的个体总数和被海刺猬用于遮蔽的贝壳总数以及两者到达稳定状态的时间,虽然有一定的差异,但都没有显著性的差异。这也进一步证明了海胆的遮蔽行为是长期进化形成的一种比较稳定的行为,短期的缺失并不会使海胆丢失这种行为,同时也说明了遮蔽物本身并不是海刺猬遮蔽行为的进化压力。遮蔽组的海刺猬,其前3个海刺猬分别初次遮蔽自己的耗时要显著短于空白组。这很可能是由于长期生活在以贝壳作为遮蔽环境的海刺猬受光照的影响更小,对光的敏感性更高,因为实验发现遮蔽组和掩蔽之间前3个海刺猬分别初次遮蔽自己的耗时并没有显著的差异。通过以上3组实验的结果比较我们推测,海刺猬的遮蔽行为只是海刺猬进化过程中形成的多种避光策略中的一种,光照有可能是该种行为的进化压力之一,而遮蔽物本身可能并不作为海刺猬形成该行为的进化压力,海刺猬最终选择何种避光策略可能跟它们生活环境中所能提供的避光条件有关。Travisano和Ratcliff通过在实验室条件下实现对酵母菌(Saccharomyces cerevisiae)由单细胞向多细胞进化以及Bell将单胞绿藻(Chlamydomonas)由光能自养型向化能自养型的演变都证明,给予生物体特定的生活条件即正确的选择压,经过大量代数的演变和积累,能够形成适应相应生活环境的生物特性[20]。马瑞俊[21]、赵彩云[22]归纳总结了关于全球气候变化对野生动物和蝴蝶的影响的研究,表明长期气候环境的改变会在一定程度上改变动物的行为和形态等生物特征。若光照刺激和生活环境中避光条件的定向改变的时间长到足以使海刺猬进行大量代数的演化与累积,就有可能会形成具有不同避光策略的海刺猬。但是由于大多多细胞生物无法像单细胞生物那样快速繁殖,而且长期定向的环境条件也不好控制,因此,对多细胞生物进行实验室条件下的演化研究还存在困难,需进一步探讨。
海胆遮蔽行为所利用的遮蔽材料是有选择性的,这可能跟遮蔽物自身的材质、规格、色彩等因素有关,也可能跟海胆自身的偏好有关[2-3,6,17]。Crook[6]发现拟球海胆只选择单一遮蔽物的比例超过59%,并且偏向选择面积体重比较大的遮蔽物,以此来降低遮蔽行为对身体能量的消耗。Sigg等[23]也发现短刺海胆倾向于选择质量更轻和更熟悉的遮蔽材料。单独以贻贝贝壳或者菲律宾蛤仔贝壳作为遮蔽物的海刺猬要显著的多于同时以两种贝壳作为遮蔽物的海刺猬,占到了78.57%,也存在对遮蔽材料的选择性。这和Crook的结果类似,进一步证明了海胆对遮蔽物的选择性。但是,与Crook和Sigg不一样的是,以菲律宾蛤仔的贝壳作为遮蔽物的海刺猬个体数要显著多于以贻贝贝壳作为遮蔽物的海刺猬个体数,菲律宾蛤仔贝壳的重量和面积要远大于贻贝贝壳。一个原因可能是由于海刺猬在其可承受的遮蔽物重量范围内,更倾向于选择面积更大的遮蔽物;另一个原因可能是在自然状态下菲律宾蛤仔白色的贝壳相对于贻贝黑色的贝壳,更利于海刺猬的伪装,减少被天敌捕食的机会。
综上所述,作为长期进化过程中保留下来的一种行为,海刺猬(G.crenularis)的遮蔽行为受光照的影响较大,光照很有可能是该行为的进化压力之一。遮蔽行为很可能是海刺猬避光策略中的一种选择。生活环境中一段时间内遮蔽物的缺失并不会使海刺猬丧失这种行为,不过会在一定程度上影响其遮蔽行为的强度和灵敏度。海刺猬对于遮蔽物的种类具有选择性,这种选择性受遮蔽物的材质、生活环境及海胆自身生理条件的影响。影响海刺猬遮蔽行为是多种因素综合作用的结果,有必要进一步研究这些因素的综合效应以及遮蔽行为对海刺猬自身生理的影响。
[1] Dumont C P,Drolet D,Deschênes I,Himmelman J H.Multiple factors explain the covering behaviour in the green sea urchin,Strongylocentrotus droebachiensis.Animal Behaviour,2007,73(6):979-986.
[2] Amato K R,Emel S L,Lindgren C A,Sullan K M,Wright P R,Gilbert J J.Covering behavior of two co-occurring Jamaican sea urchins:differences in the amount of covering and selection of covering material.Bulletin of Marine Science,2008,82(2):255-261.
[3] Verling E,Crook A,Barnes D.Covering behaviour in Paracentrotus lividus:is light important?Marine Biology,2002,140(2):391-396.
[4] Dix T G.Covering response of the echinoid Evechinus chloroticus(Val.).Pacific Science,1970,24(2):187-194.
[5] James D W.Diet,movement,and covering behavior of the sea urchin Toxopneustes roseus in rhodolith beds in the Gulf of California,México.Marine Biology,2000,137(5/6):913-923.
[6] Crook A C.Individual variation in the covering behaviour of the shallow water sea urchin Paracentrotus lividus.Marine Ecology,2003,24(4):275-287.
[7] Barnes D,Crook A.Quantifying behavioural determinants of the coastal European sea-urchin Paracentrotus lividus.Marine biology,2001,138(6):1205-1212.
[8] Verling E,Crook A C,Barnes D K A.The dynamics of covering behaviour in dominant echinoid populations from American and European west coasts.Marine Ecology,2004,25(3):191-206.
[9] Kehas A J,Theoharides K A,Gilbert J J.Effect of sunlight intensity and albinism on the covering response of the Caribbean sea urchin Tripneustes ventricosus.Marine Biology,2005,146(6):1111-1117.
[10] Millott N.The covering reaction of sea-urchins:1.A preliminary account of covering in the tropical echinoid Lytechinus variegatus(Lamarck),and its relation to light.Journal of Experimental Biology,1956,33:508-523.
[11] Millott N.The photosensitivity of echinoids.Advances in Marine Biology,1976,13:1-52.
[12] Lees D C,Carter G A.The covering response to surge,sunlight,and ultraviolet light in Lytechinus anamesus(Echinoidea).Ecology,1972,53(6):1127-1133.
[13] Richner H,Milinski M.On the functional significance of masking behaviour in sea urchins-an experiment with Paracentrotus lividus.Marine Ecology Progress Series,2000,205:307-308.
[14] Crook A C,Barnes D K A.Seasonal variation in the covering behaviour of the echinoid Paracentrotus lividus(Lamarck).Marine Ecology,2001,22(3):231-239.
[15] Chang Y Q,Ding J,Song J,Yang W.Biology Research and Breeding of Sea Cucumber and Sea Urchin.Beijing:Ocean Press,2004:220-221.
[16] Luo S B,Chang Y Q,Zhao C,Zhou H S.Effects of the covering behavior on food consumption,growth and gonad traits of the sea urchin Glyptocidaris crenularis.Acta Ecologica Sinica,2012,doi:10.5846/stxb201111241797.
[17] Adams N L.UV radiation evokes negative phototaxis and covering behavior in the sea urchin Strongylocentrotus droebachiensis.Marine Ecology Progress Series,2001,213:87-95.
[18] Amsler C D,McClintock J B,Baker B J.An Antarctic feeding triangle:defensive interactions between macroalgae,sea urchins,and sea anemones.Marine Ecology,1999,183:105-114.
[19] Sharp D T,Gray I E.Studies on factors affecting the local distribution of two sea urchins,Arbacia punctulata and Lytechinus variegatus.Ecology,1962,43(2):309-313.
[20] Pennisi E.Evolutionary time travel.Evolutionary Biology,2011,334(6058):893-895.
[21] Ma R J,Jiang Z G.Impact of global climate change on wild life.Acta Ecologica Sinica,2005,25(11):3061-3066.
[22] Zhao C Y,Li J S,Luo J W,Xiao N W,Luo Z L.A review on responses of butterflies to global climate chang.Acta Ecologica Sinica,2010,30(4):1050-1057.
[23] Sigg J E,Lloyd-Knight K M,Boal J G.UV radiation influences covering behaviour in the urchin Lytechinus variegatus.Journal of the Marine Biological Association of the United Kingdom,2007,87(5):1257-1261.
参考文献:
[15] 常亚青,丁君,宋坚,杨威.海参、海胆生物学研究与养殖.北京:海洋出版社,2004:220-221.
[16] 罗世滨,常亚青,赵冲,周海森.遮蔽行为对海刺猬(Glyptocidaris crenularis)摄食、生长和性腺性状的影响.生态学报,2012,DOI:10.5846/stxb201111241797.
[21] 马瑞俊,蒋志刚.全球气候变化对野生动物的影响.生态学报,2005,25(11):3061-3066.
[22] 赵彩云,李俊生,罗建武,肖能文,罗遵兰.蝴蝶对全球气候变化响应的研究综述.生态学报,2010,30(4):1050-1057.
猜你喜欢
检察风云(2024年10期)2024-05-24 17:38:09
食品与生活(2024年3期)2024-03-19 08:59:32
渔业现代化(2023年2期)2023-04-23 01:10:18
小猕猴学习画刊(2022年10期)2022-11-01 05:35:32
新农业(2022年6期)2022-04-13 13:37:47
幼儿画刊(2021年9期)2021-09-20 11:35:42
烹调知识(2020年10期)2020-10-20 05:47:44
水产养殖(2016年2期)2016-03-28 11:47:52
浙江大学学报(农业与生命科学版)(2015年4期)2015-12-15 12:47:42
餐饮世界(2015年6期)2015-08-07 03:08:50
