
不同干扰对黄土区典型草原物种多样性和生物量的影响
2013-12-16 08:18:52陈芙蓉程积民朱仁斌杨晓梅赵新宇苏纪帅
生态学报 2013年9期
陈芙蓉,程积民,刘 伟,朱仁斌,杨晓梅,赵新宇,苏纪帅
(1.中国科学院水利部水土保持研究所,杨凌 712100;2.中国科学院研究生院,北京 100049;3.国家林业局中南林业调查规划设计院,长沙 410014;4.西北农林科技大学,杨凌 712100)
典型草原是在温带内陆半干旱气候条件下形成的草原类型,以旱生的多年生丛生禾草占优势的草原植被,并以针茅属的物种为其主要的物种成分[1]。云雾山自然保护区是我国黄土高原半干旱地区干草原生态系统的典型地段,同时也是以本氏针茅(Stipa bungeana)为建群种的干草原生态系统保留较完整、面积最大、原生性最强的典型地区,代表着黄土高原半干旱草原的自然特征[2]。草地是受人类活动影响最大的陆地生态系统之一,研究不同干扰对其生物多样性和生物量的影响是草地生态系统恢复与重建工作中的重要内容[3]。近些年来针对黄土区半干旱典型草地退化的问题,云雾山自然保护区主要采取围栏封育措施来恢复重建,因此已有很多关于封育对草地植被恢复方面的研究,提出封育可以显著影响草地群落特征,提高退化草地的生产力[4-5]。此外,放牧、施肥和火烧是人类在草地生态系统管理实践中施加于草地的主要干扰类型,对生态系统过程会产生重要的影响。如施肥使高寒草甸植物群落物种组成贫乏,群落结构趋于简单,物种多样性降低[6-8]。放牧通过改变植被结构和土壤理化性质,增加了生境的异质性,从而造成草地群落植物组分、结构和多样性格局发生变化,进而对整个生态系统的结构和功能产生影响[6,9]。国外对草原火烧的研究比较广泛,不仅关注草原火对不同植物种类的生长发育和群落生产力的影响,而且还把火烧作为一种草原管理工具进行研究,探讨火烧对抑制和消除草原灌木、提高草场的利用价值等方面的作用[10-11]。本文主要通过野外调查研究施肥、封育、火烧和放牧4种不同的干扰类型对黄土区典型草原植被群落物种组成、多样性和地上生物量变化的影响,以期为草原生物多样性的保护和草原的可持续利用管理提供科学依据。
1 材料与方法
1.1 研究区概况
研究区位于宁夏云雾山国家级草原自然保护区,它是我国黄土高原半干旱区最完整、面积最大的典型草原生态系统,代表了黄土高原半干旱区的自然特征。地理位置106°24'—106°28'E,36°13'—36°19'N,面积6700 hm2,海拔1 800—2 100 m,年平均气温5℃,最热月为7月,气温在22—25℃,最冷月为1月,平均最低气温-14℃,≥0℃积温2370—2 882℃,年日照2500 h,无霜期112—140 d。年降水量400—450 mm,降水季节分配不均,全年65%—75%的降水集中在7—9月,年蒸发量1 330—1 640 mm。地势南低北高,阳坡平缓,阴坡较陡,属长梁缓坡丘陵区,土层深厚,土壤类型为淡黑垆土和山地灰褐土两类。该区自1982年开始封育,植被主要是温带典型草原,主要草本植物有本氏针茅、百里香(Thymus mongolicus)、铁杆蒿(Artemisia sacrorum)、大针茅(S.grandis)、白颖苔草 (Carex rigesaens)、星毛委陵菜(Comarum acaulis)、赖草(Leymus secalinus)等,群落自然恢复演替序列为冰草—百里香+杂类草—本氏针茅+百里香—本氏针茅+铁杆蒿—本氏针茅+大针茅[12],目前丛生禾本科植物本氏针茅在该区分布范围最广[2]。
1.2 实验设计
在云雾山草原自然保护区内天然草地上设置不同干扰类型的样地。2011年5月共设置4个地形一致、水热条件相同的阳坡样地:施肥样地、火烧样地、放牧样地和封育样地,由于施肥样地和火烧样地都设置在封育区内,所以按不同干扰方式设置4个处理,分别为封育+施肥、封育+火烧、封育和放牧,样地具体情况如表1。
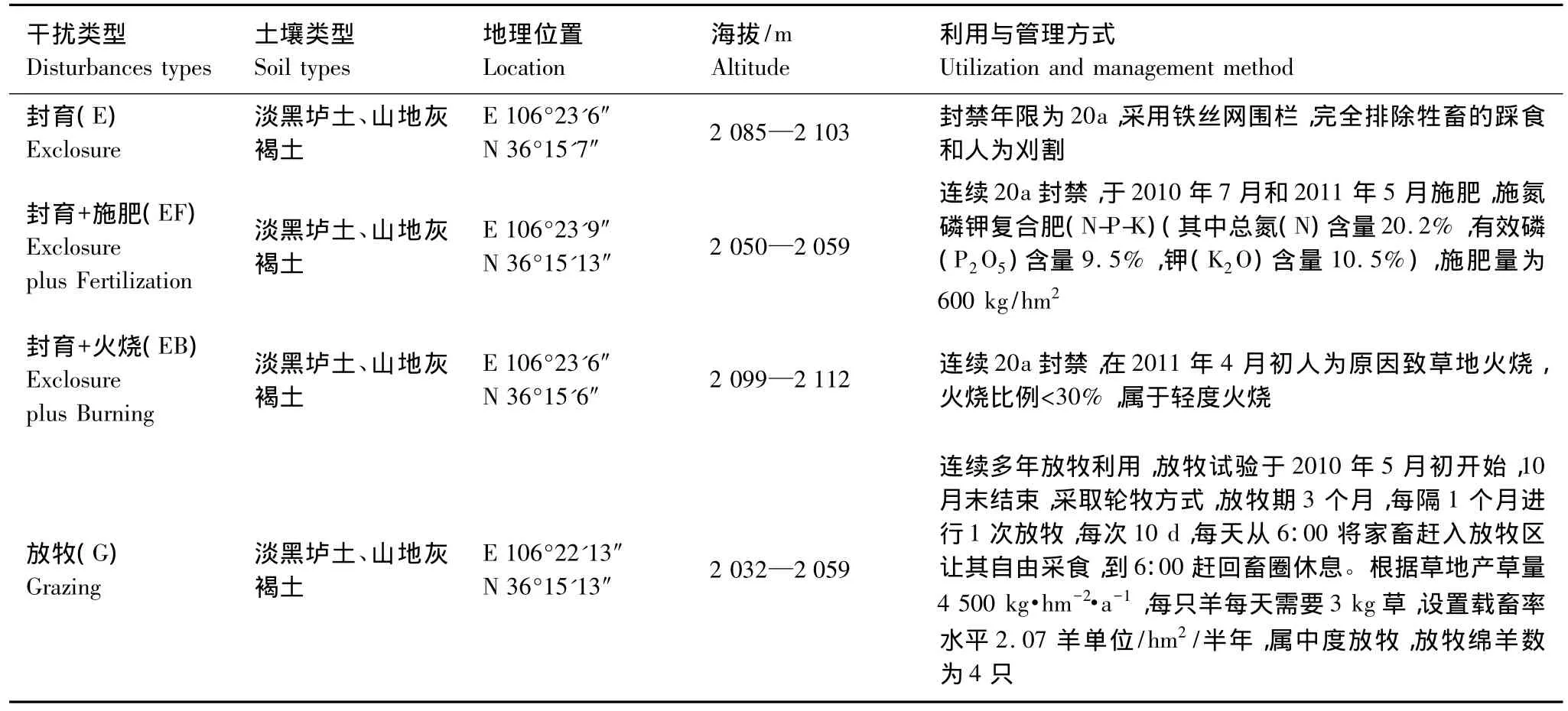
表1 调查样地的基本信息Table 1 Basic information of sampled plots
1.3 研究方法
调查方法:调查时间为2011年9月份,考虑到地形因素的影响,在每个样地选取坡底、坡中和坡上3个坡位,在每个坡位随机选取5个样方(50 cm×50 cm),调查每个样方中出现的物种并测量高度、盖度、多度等,记录优势种。
环境因子测定:用GPS测定每个样地的高程和地理坐标,用温湿表测定气温和空气湿度。
植物样调查:采用针刺法测定样方总盖度和每种植物的分盖度;每种植物随机选5株,直尺测量其伸长高度;对样方中出现的植物进行分类,记录同一种植物出现的个数,并将其地上部分分种齐地面刈割,除去黏附的土壤、砾石、杂物后装袋,然后收集样方中所有凋落物,带回实验室然在(65±2)℃条件下烘干至恒重,称其干重,地上生物量和凋落物量均采用干重计算。
1.4 计算方法
(1)群落内各物种重要值的计算[13]
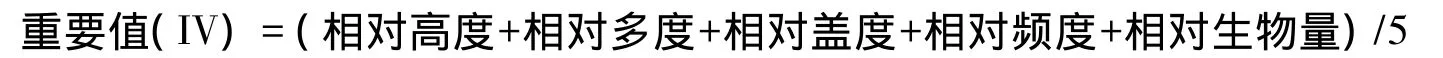
(2)地上现存量
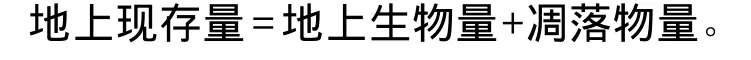
(3)物种多样性分析
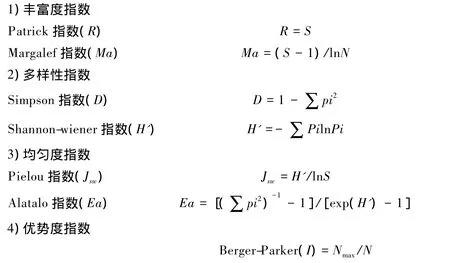
式中,S为样地中的物种数,Pi为样地中第i种的生物量占全部生物量的比例,Nmax为群落中最大种的生物量,N 为群落的总生物量[8]。
1.5 数据分析
采用Excel软件对数据进行预处理,采用SAS V8统计软件对数据进行统计分析。由Shapiro-Wilk检验发现数据符合正态分布,采用ANOVA分析方法检验各处理在P=0.05显著水平上的差异。如差异显著,采用Duncan法在P=0.05显著水平上进行多重比较。采用Pearson法分析多样性和生产力的相关性。
2 结果
2.1 不同干扰对群落物种组成和结构的影响
不同干扰样地的优势种不同,反映出不同的群落外貌特征。如表2所示,4种不同干扰样地共出现45种植物,分属23个科,优势种主要出现在禾本科、莎草科、豆科和菊科。其中,施肥地有30种,重要值最高的是草甸草原的指示植物莎草科白颖苔草,其次是本氏针茅和大针茅;火烧地出现植物28种,重要值最高的是铁杆蒿,因其抗旱能力和根蘖性强,火烧后成为群落的优势种,其次为本氏针茅和大针茅;封育地有植物26种,大针茅、硬质早熟禾和花苜蓿占优势地位;放牧地出现的植物种类最多,有33种,优势种为耐干旱的扁穗冰草和耐践踏的星毛委陵菜。
植物群落结构可以通过其高度、总盖度和密度加以反映[14]。不同干扰的4个样地的群落盖度为封育+施肥>封育+火烧>封育>放牧地,分别为89%,76%,72%和53%,其中封育+施肥样地的群落盖度显著高于其他3种干扰样地(P<0.05),放牧地的群落盖度显著低于其他3种干扰样地(P<0.05)(图1)。封育+施肥、封育+火烧、封育和放牧样地的群落高度分别为23.57,22.60,22.41和15.58 cm,其中放牧地的群落高度显著低于其他3种干扰样地(P<0.05),封育+施肥、封育+火烧和封育3种干扰样地之间没有明显差异(图1)。不同干扰样地的群落密度为封育(1 111.7株/m2)>封育+施肥(1 019.7株/m2)>封育+火烧(889.1株/m2)>放牧(693.9株/m2),其中,封育与封育+施肥样地显著大于放牧地(P<0.05)(图1)。4种干扰类型中,封育地的密度最大,主要是群落中豆科花苜蓿和杂类草阿尔泰狗娃花的贡献所致。
2.2 不同干扰对群落地上现存量的影响
如图图2,不同干扰的样地地上生物量为封育+施肥>封育+火烧>封育>放牧地,分别为367.78,312.31,255.46和172.82 g/m2,其中封育+施肥草地的地上生物量最高,显著高于其他3种干扰方式(P<0.05),放牧地地上生物量最低,显著低于其他3种干扰方式(P<0.05),且不同干扰方式之间均有显著差异(P<0.05),可见施肥显著提高了群落的初级生产力。从枯落物量来看,封育+施肥和封育样地显著高于放牧样地(P<0.05),由于火烧使样地枯落物为0,所以显著低于其他3种干扰方式(P<0.05)。从地上现存量(地上生物量+枯落物量)来看,封育+施肥样地的地上现存量最高,为545.71 g/m2,显著高于其他3种干扰方式(P<0.05);封育+火烧与封育样地没有明显差异;放牧地的地上现存量最低,为212.29 g/m2,显著低于其他3种干扰方式(P<0.05),可见放牧限制群落生产力的发挥。
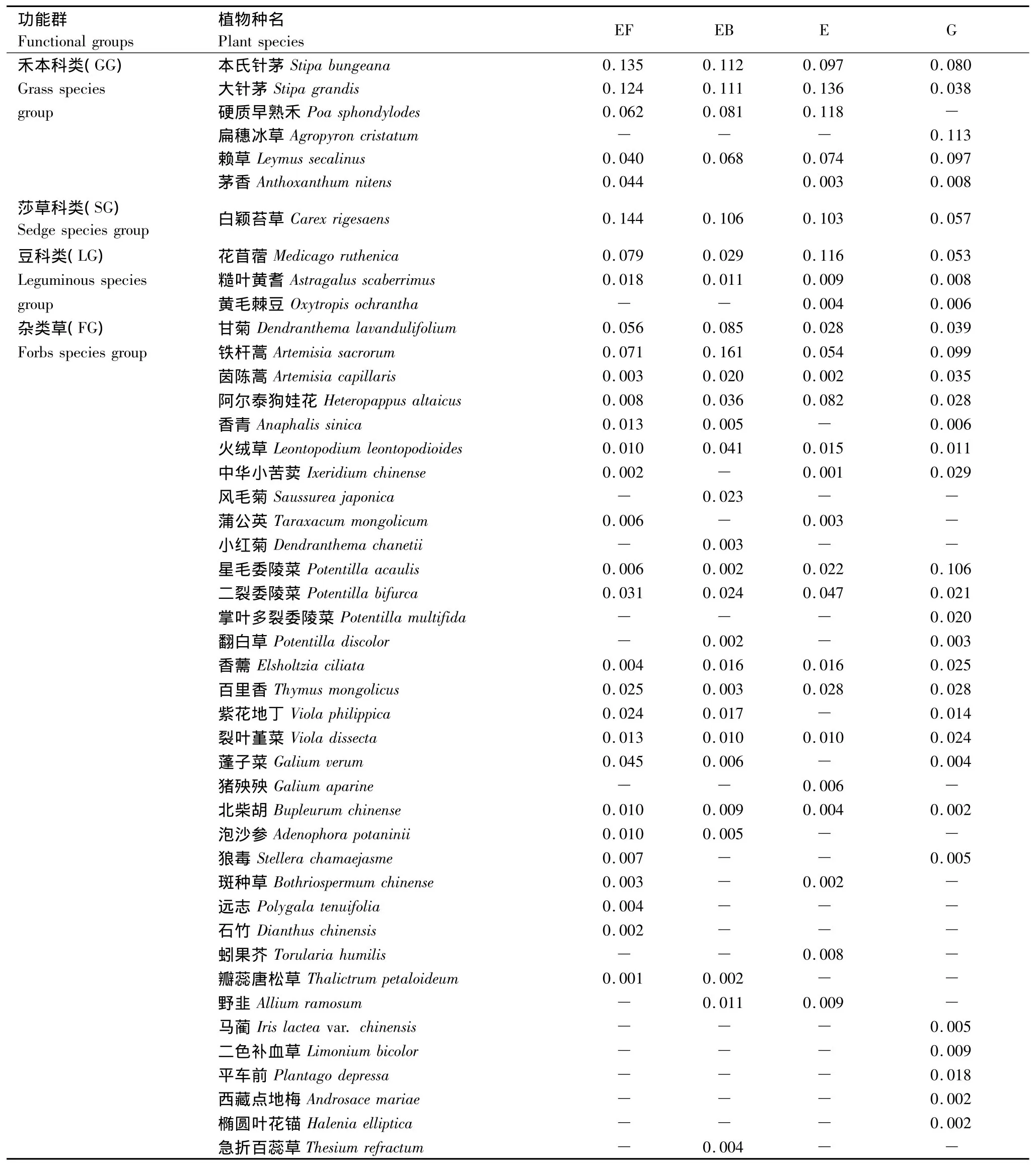
表2 不同干扰下典型草原群落物种组成和重要值Table 2 Species composition and important values of the community under different disturbances in typical steppe
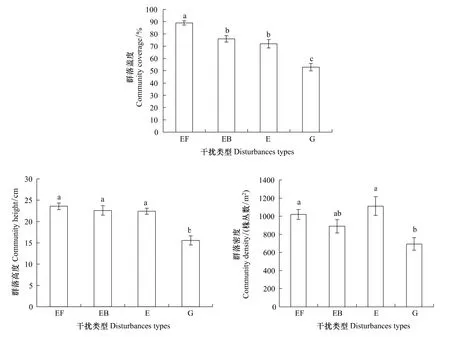
图1 不同干扰下典型草原群落的外貌特征Fig.1 Community features of different disturbances in typical steppe
从生活型功能群来看(图2),不同干扰样地各功能群类型基本一致,均由禾本科类、莎草类、豆科类和杂类草组成,但各功能群生物量在群落中所占的比例则大不相同,禾本科草与杂类草约占85%—94%。在封育+施肥样地与封育样地,禾本科草生物量显著高于杂类草,占60%左右,处于优势地位。而封育+火烧与放牧地,禾本科草与杂类草所占整个群落的比例大致相同,在45%左右。在4种干扰类型中,封育+施肥地的地上生物量最高,主要是禾本科草类与莎草类显著高于其他3种干扰类型所致(P<0.05)。4种干扰类型的豆科类与杂类草生物量没有显著差异。
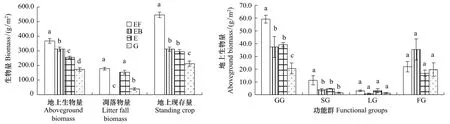
图2 不同干扰对典型草原群落地上生物量的影响Fig.2 Effects of different disturbances on above-ground biomass in typical steppe
2.3 不同干扰对群落物种多样性的影响
在不同干扰条件下群落物种多样性的变化不同(图3)。物种丰富度指数是表明群落中物种多寡的参数,丰富度指数Ma指标表明放牧地群落物种丰富度最高,为4.943,封育地物种丰富度最低,为3.642;不同干扰群落丰富度指数(R和Ma)为放牧>封育+施肥>封育+火烧>封育。均匀度反映各群落中物种个体数分布的均匀程度,均匀度指数(Jsw和Ea)指标都反映出封育样地群落均匀度指数最高,封育+施肥样地群落均匀度指数最低,封育+火烧与放牧样地居中。多样性指数是物种水平上多样性和异质性程度的度量,综合反映群落物种丰富度和均匀度。所以它与物种丰富度和均匀度的结果有一定差异。不同干扰样地群落多样性指数(D和H′)为放牧>封育>封育+火烧>封育+施肥。优势度指数(DB)所表达的信息与多样性指数和均匀度指数相反,反映了各物种种群数量的变化情况,不同干扰群落优势度指数(DB)为封育+施肥>封育+火烧>封育>放牧,封育+施肥样地优势度指数最大,为0.263,说明群落内物种数量分布最不均匀,优势种大针茅的地位突出;封育+火烧地次之,这与铁杆蒿在群落中分布多而集中占优势有关(图3)。
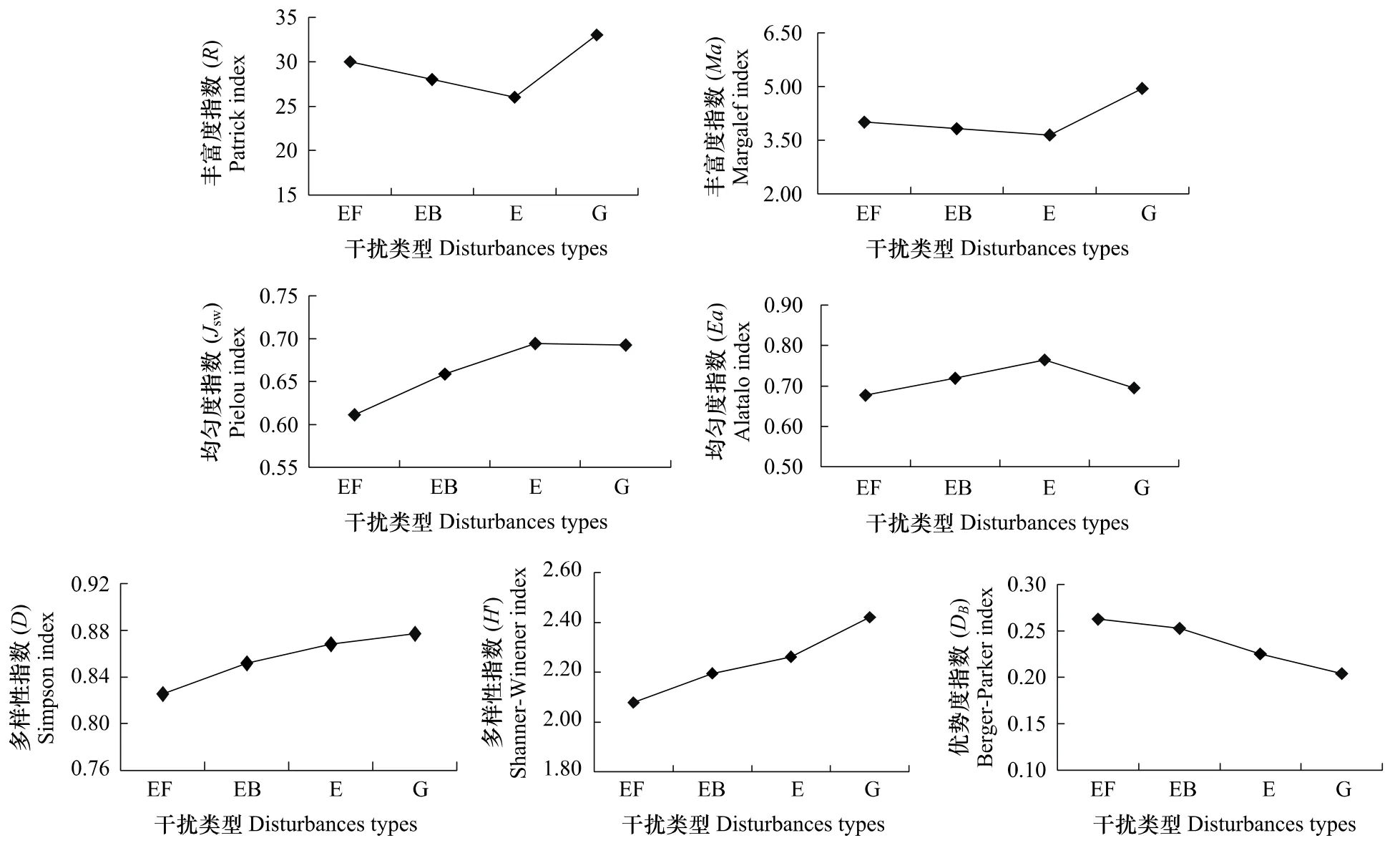
图3 不同干扰对典型草原群落物种多样性的影响Fig.3 Effects of the different disturbances on the diversity index of community in typical steppe
2.4 群落生产力与多样性指数的关系
群落物种多样性是丰富度和均匀度的综合,它又是判断生物群落结构变化或生态系统稳定性的指标,本文通过对多样性(Shannon-Wiener和Simpson)指数和地上生物量的相关分析表明:地上生物量与多样性之间呈显著负相关(图4)。
3 讨论与结论
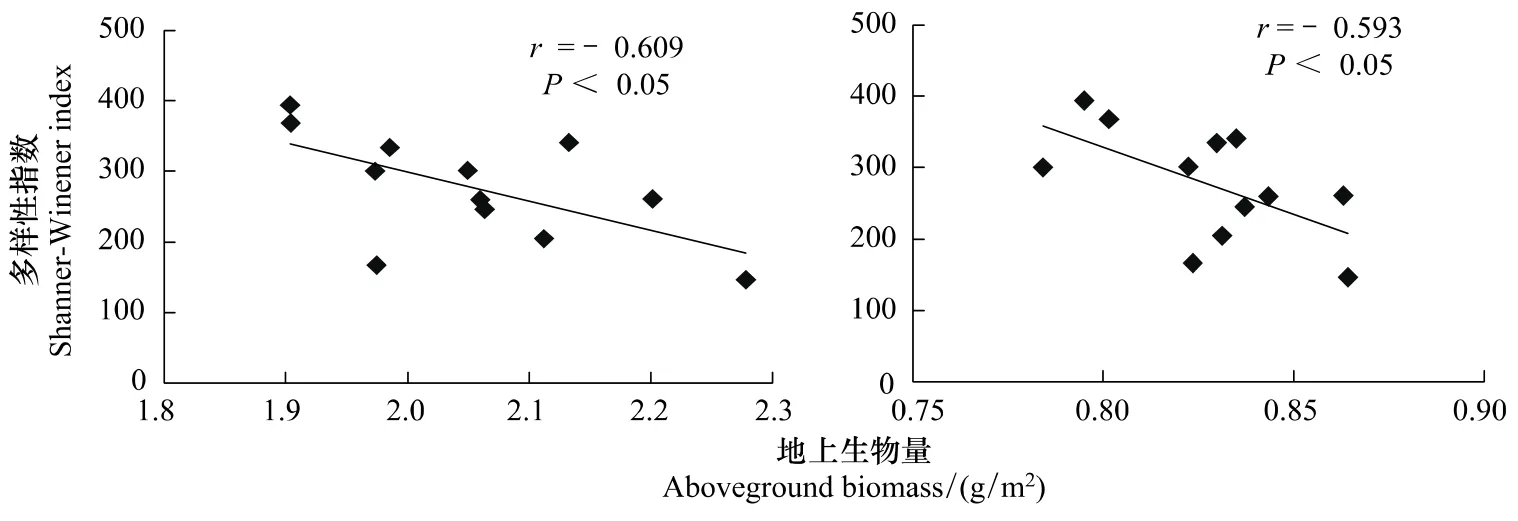
图4 典型草原地上生物量与物种多样性的关系Fig.4 The relationships between above-ground biomass and diversity index in typical steppe
放牧是草地常见的干扰方式,也是一种高度复杂的干扰方式,它对植物群落既有消极作用,也有积极作用[15]。本研究结果表明,放牧与其他干扰方式相比,样地群落高度、盖度、密度和地上生物量显著降低,这与前人研究结果一致[16-17]。因为牲畜的采食具有选择性,加上采食过程中的踩踏,使得牲畜喜食牧草和不耐践踏牧草数量急剧减少,进而改变草地群落结构,影响草地生产力的积累。另一方面放牧也是草地生态系统生物多样性变化的重要原因之一[18-19]。不同的放牧强度对植物多样性的影响程度不同,研究表明[9]适度放牧能显著提高草地群落的物种多样性,本实验放牧样地属于中度放牧,丰富度指数和多样性指数都高于其他干扰方式,符合中度干扰理论[3,20]:即中等干扰条件下,物种多样性最大。优势度指数最小,均匀度指数低于封育样地,说明放牧增加了群落的空间异质性。因为首先牲畜的啃食抑制了优势种本氏针茅的生长,一些耐牧草种(扁穗冰草、铁杆蒿等)、耐践踏草种(星毛委陵菜)和牲畜不喜食的杂类草明显增加,即降低了优势种的竞争优势,所以优势度指数低于其他3种干扰样地,这样为竞争力较弱物种的生存拓宽了空间,提高了资源利用率,使物种在种类和数量配置上发生变化,增加了群落结构的复杂性,导致生物多样性发生变化[8]。其次家畜在采食过程中可以帮助某些植物传播种子,扩大了一些植物的分布面积[21]。
相对于放牧干扰,围栏封育是退化草地恢复与重建的重要措施之一,对云雾山草地实施封育后,草地植被盖度、平均高度以及地上、地下生物量,均有不同程度的提高[22-23]。本研究结果表明:封育和放牧样地的植被盖度、高度和地上现存量的差异均达到显著水平(P<0.05)。王炜[24]认为草原退化群落在自然封育条件下迅速恢复的原因是植物在消除放牧干扰后种群拓殖与对群落剩余资源利用的能力增加了。但封育对群落多样性影响的研究结果较复杂[7,25],贾晓妮[4]研究得出围栏封育在一定时间范围内可以显著改善群落的特征,增加群落的生产力,但围封年限不同会显著影响物种多样性,指出了封育最佳年限为10—15a。本实验封育样地年限为20a,群落水平结构均一,垂直结构从高到低依次为大针茅、硬质早熟禾和多年生牧草花苜蓿,而且其重要值都高于其他3种干扰样地,这与邹厚远等[10]对黄土高原典型草原的自然恢复演替推断的第5个阶段相似,说明退化草地经过内因生态演替达到与当地气候相对稳定的群落。封育样地物种丰富度指数低于其他3种干扰样地,物种多样性指数低于放牧地,可见长时间的封育会减少植物物种多样性,与前人研究结果一致[26-27],但物种均匀度指数高于其他3种样地,群落空间匀质性高。根据草地生态系统的可持续性原理,草地围封不应是无限期的。围栏封育时间的长短应根据草地退化程度和草地恢复状况而定,封育期过长,枯落物积累太多会抑制植物的再生和幼苗的形成,不利于草地的繁殖更新,而且会降低草地物种多样性,影响草地生态系统的能量流动和物质循环,对草地健康会产生严重干扰。
对封育20a的样地进行火烧后,群落盖度、高度和密度没有显著变化,但地上生物量显著增加(P<0.05),所以火烧能显著增加群落生产力[28]。首先本研究样地在生长季初期遭火烧,消除了地上立枯物和凋落物,为竞争力较弱的物种的进入提供了生存空间,多年生植物和杂类草种类(急折百蕊草、瓣蕊唐松草、翻白草等)增加;其次由于地表覆盖度减小,地表接受到的光照强度和土壤温度的增加可刺激种子萌发[29],所以物种丰富度指数高于封育样地,这与Collins[30]研究早春火烧使北美混合草原物种丰富度增加的结果一致。但地表长时间的裸露,使土壤水分损失较多,对植物个体发育和群落发展有不利影响,所以封育+火烧样地群落密度与放牧地相近,且多样性指数和均匀度指数低于封育样地。这与李晓波[31]、贺郝钰[32]等研究结果一致。此外针茅属于根茎型禾草,耐火烧能力强[33],在草地返青季,他们能很快生长,成为群落优势种,可见火烧后本氏针茅和大针茅的优势种地位没有变化,不同的是铁杆蒿是一种半灌木,根蘖性强,生命力也极强,在火烧后迅速分蘖出枝条,而且铁杆蒿还能分泌出一种化感物质,抑制其他物种的生长[34],成为群落的建群种,因此封育20a的草地火烧后,群落有向铁杆蒿+本氏针茅+大针茅阶段逆向演替的迹象,但这个结论还有待进一步研究。
对封育20a的草地进行施肥后,群落盖度和地上现存量显著高于其他3种干扰方式(P<0.05),因为施肥增加了土壤中的养分资源,一些喜肥的物种(本氏针茅和大针茅)具有比其它物种更快的生长率,促进自身地上生物量和盖度的增加,最终引起群落生产力的增加[35]。而物种丰富度指数高于封育和封育+火烧地,这与前人研究结果不同[36],原因可能是施肥年限少,植物种类影响还不明显。但群落物种多样性指数和均匀度指数都低于其他干扰样地,这与前人研究结果一致[7-8]。李伟[37]认为光竞争假说和根竞争假说都不能全面解释施肥后多样性降低,地上/地下竞争的相互作用可能是施肥后多样性降低的最合理的解释。因为随着群落中优势种的旺盛生长,群落的密闭度增加,自身光资源截取和光竞争的强度增加,使群落下层不耐荫物种或个体较小物种在群落中减少或死亡。同时,资源的增加改变了群落内物种的相对竞争能力,使草地中优势种(针茅类)地下、地上竞争能力增强,从而排除了杂草和豆科物种,使多样性降低[38]。可见封育草地施肥后多样性下降符合生态位理论[37],即生态位相同的物种因竞争相同的资源而不能稳定共存。另外,由于施肥后白颖苔草的相对密度增加导致其重要值最高,可推测为这是黄土高原典型草原在外因作用下的演替结果,即白颖苔草取代了本氏针茅和大针茅的优势地位,使群落发展成为更加典型的草甸群落。
本实验中,地上生物量与物种多样性呈负相关关系,不支持多样性-生产力假说(即建立在不同物种利用不同资源的基础上,复杂多样的植物群落能利用更多的有限资源从而获得更高的生产力[39]),而更倾向于竞争假说(根竞争和光竞争),Rajaniemi[40]认为,在生产力低且资源斑块化的植物群落中,根竞争成为个体大小不对称竞争,导致物种多样性下降。在高生产力条件下,物种间竞争由地下转为地上,随着生产力的提高,长得高的物种将遮住矮的物种,这种对光的大小不对称竞争将导致更多的物种被排除。在不平衡的扰动状态中,根竞争的作用可能特别重要;但在平衡的生产力很高的群落中,光竞争会对物种多样性与生产力的关系起重要作用,所以地上/地下竞争的相互作用共同导致了生产力与多样性的负相关关系。
[1] Chen Z Z,Wang S P.Typical Grassland Ecosystem in China.Beijing:Science Press,2000:1-6.
[2] Management office of Yunwu Mountain natural conservation area in Ningxia.Collected works of scientific investigation and management in Yunwu Mountain natural conservation area in Ningxia.Yinchuan:Ningxia People's Press,2001.1-131.
[3] Wen L,Dong S K,Zhu L,Shi J J,Liu D M,Wang Y L,Ma Y S.The effect of natural factors and disturbance intensity on spacial heterogeneity of plant diversity in alpine meadow.Acta Ecologica Sinica,2011,31(7):1844-1854.
[4] Jia X N,Cheng J M,Wan H E.Change of species diversity on typical Stipa bungeana community restoration and succession in the Yunwu Mountain.Acta Prataculturae Sinica,2008,17(4):12-18.
[5] Cheng J M,Wan H E,Hu X M.Study of vegetation restoration and rebuilding pattern and the process of succession in the loess hilly regions.Acta Agrestia Sinica,2005,13(4):324-327.
[6] Proulx M,Mazumder A.Reversal of grazing impact on plant species richness in nutrient-poor vs.nutrient-rich ecosystems.Ecology,1998,79(8):2581-2592.
[7] Zhou G Y,Chen G C,Zhao Y L,Wang S Z,Li W,Peng M.Comparative research on the influence of chemical fertilizer application and enclosure on alpine steppes in the Qinghai Lake area Ⅰ.Structure and species diversity of the plant community.Acta Pratacultural Science,2004,13(1):26-31.
[8] Jiang X L,Zhang W G,Yang Z Y,Wang G.The influence of disturbance on community structure and plant diversity of alpine meadow.Acta Botanica Boreali-occidentalia Sinica,2003,23(9):1479-1485.
[9] Yang L M,Han M,Li J D.Plant diversity change in grassland communities along a grazing disturbance gradient in the northeast China transect.Acta Phytoecologica Sinica,2001,25(1):110-114.
[10] Hensel R L.Recent studies on the effect of burning on grassland vegetation.Ecology,1923,4(2):183-188.
[11] Anderson H G,Bailey A W.Effects of annual burning on grassland in the aspen parkland of east-central Alberta.Canadian Journal of Botany,1980,58(8):985-996.
[12] Zou H Y,Cheng J M,Zhou L,Hongo A.Natural recoverage succession and regulation of the prairie vegetation on the loess plateau.Research of Soil and Water Conservation,1998,5(1):127-138.
[13] Li R,Zhang K B,Liu Y F,Wang B T,Yang X H,Hou R P.Plant community spatial distribution pattern of wetland ecosystem in a semi-arid region of northwestern China.Journal of Beijing Forestry University,2008,30(1):6-13.
[14] Ren J Z.Research methods for pratacultural science.Beijing:China Agriculture Press,1998:1-30.
[15] Mclntyre S,Lavorel S,Landsberg J,Forbes T D A.Disturbance response in vegetation-towards a global perspective on functional traits.Journal of Vegetation Science,1999,10(5):621-630.
[16] Belsky A J.Effects of grazing,competition,disturbance and fire on species composition and diversity in grassland communities.Journal of Vegetation Science,1992,3(2):187-200.
[17] Pettit N E,Froend R H,Ladd P G.Grazing in remnant woodland vegetation:changes in species composition and life form groups.Journal of Vegetation Science,1995,6(1):121-130.
[18] Sala O E,Oesterheld M,León R J C,Soriano A.Grazing effects upon plant community structure in subhumid grasslands of Argentina.Vegetatio,1986,67(1):27-32.
[19] Austin M P,Williams O B,Belbin L.Grassland dynamics under sheep grazing in an Australian Mediterreanean type climate.Vegetatio,1981,46-47(1):201-212.
[20] Ram J,Singh J S,Singh S P.Plant biomass,species diversity and net primary production in a central Himalayan high altitude grassland.Journal of Ecology,1989,77(2):456-468.
[21] Wang R Z.Effects of disturbances on species diversity in grassland ecosystems.Journal of Northeast Normal University:Natural Science Edition,1996,(3):112-116.
[22] Jia X N,Cheng J M,Wan H E.Effects of enclosure on the community characteristics of stipa bungeana grassland in Yunwu Mountain.Acta Agrestia Sinica,2008,16(3):272-277.
[23] Chen F R,Cheng J M,Yu L N,Li Y,Wu Y Q.Effects of fencing and grazing on the biomass of typical steppe in the Loess Plateau.Pratacultural Science,2011,28(6):1079-1084.
[24] Wang W,Liu Z L,Hao D Y,Liang C Z.Research on the restoring succession of the degenerated grassland in Inner Mongolia—Ⅰ.Basic characteristics and driving force for restoration of the degenerated grassland.Acta Phytoecologica Sinica,1996,20(5):449-459.
[25] Qi B,Zhang D G,Ding L L,Zhang L,Wang X,Yang Y H.Properties of plant community diversity of degraded alpine arid grasslands.Journal of Gansu Agricultural University,2005,40(5):626-631.
[26] Fan Y G,Hu Y K,Li K H,Yu J M,Wang X.Effects of different disturbances on the diversity and biomass of the phytobiocoenoses in alpine steppes.Arid Zone Research,2008,25(4):531-536.
[27] Hobbs R J,Huenneke L F.Disturbance,diversity,and invasion:implications for conservation.Conservation Biology,1992,6(3):324-337.
[28] Pei C F,Jiao J S.The Influence of grassland fire on Stipa krylovii and Agropyron cristatum.Grassland of China,1998,(3):39-40,46-46.
[29] Knapp A K,Seastedt T R.Detritus accumulation limits productivity of tallgrass prairie.Bioscience,1986,36(10):662-668.
[30] Collins S L.Interaction of disturbances in tallgrass prairie:a field experiment.Ecology,1987,68(5):1243-1250.
[31] Li X B,Zhou D W,Jiang S C.Effects of fire factors on plant diversity of Songnen Aneurolepidium Chinensis grassland.Pratacultural Science,1997,14(4):61-64.
[32] He H Y,Su J Q,Huang L,Jia X L,Li X R.Effects of fire on the structure of herbage synusia vegetation in desertified steppe,North China.Acta Ecologica Sinica,2011,31(2):364-370.
[33] Zhou D W,Jiang S C,Hu Y J.Changes in plant height growth,moisture and chlorophyll content following a grassland burning.Journal of Northeast Normal University:Natural Science Edition,1999,(4):91-95.
[34] Wang H,Xie Y S,Yang Y L,Chuai J F.Allelopathic effect of extracts from Artemisia sacrorum leaf and stem on four dominant plants of enclosed grassland on Yunwu Mountain.Acta Ecologica Sinica,2011,31(20):6013-6021.
[35] Tilman D,Pacala S.The Maintenance of Species Richness in Plant Communities.Chicago:University of Chicago Press,1993:557-569.
[36] Niu K C,Luo Y J,Choler P,Du G Z.The role of biomass allocation strategy on diversity loss due to fertilization.Basic and Applied Ecology,2008,9(5):485-493.
[37] Li W.Research on the Mechanism of Species Diversity Loss Due to Fertilization of Alpine Meadow on the Tibetan Plateau[D].Lanzhou:Lanzhou University,2011.
[38] Gilliam F S.Response of the herbaceous layer of forest ecosystems to excess nitrogen deposition.Journal of Ecology,2006,94:1176-1191.
[39] McCann K S.The diversity-stability debate.Nature,2000,405(6783):228-233.
[40] Rajaniemi T K.Why does fertilization reduce plant species diversity?Testing three competition-based hypotheses.Journal of Ecology,2002,90(2):316-324.
参考文献:
[1] 陈佐忠,汪诗平.中国典型草原生态系统.北京:科学出版社,2000:1-6.
[2] 宁夏云雾山草原自然保护区管理处.宁夏云雾山自然保护区科学考察与管理文集.银川:宁夏人民出版社,2001.
[3] 温璐,董世魁,朱磊,施建军,刘德梅,王彦龙,马玉寿.环境因子和干扰强度对高寒草甸植物多样性空间分异的影响.生态学报,2011,31(7):1844-1854.
[4] 贾晓妮,程积民,万惠娥.云雾山本氏针茅草地群落恢复演替过程中的物种多样性变化动态.草业学报,2008,17(4):12-18.
[5] 程积民,万惠娥,胡相明.黄土丘陵区植被恢复重建模式与演替过程研究.草地学报,2005,13(4):324-327.
[7] 周国英,陈桂琛,赵以莲,王顺忠,李伟,彭敏.施肥和围栏封育对青海湖地区高寒草原影响的比较研究Ⅰ群落结构及其物种多样性.草业学报,2004,13(1):26-31.
[8] 江小蕾,张卫国,杨振宇,王刚.不同干扰类型对高寒草甸群落结构和植物多样性的影响.西北植物学报,2003,23(9):1479-1485.
[9] 杨利民,韩梅,李建东.中国东北样带草地群落放牧干扰植物多样性的变化.植物生态学报,2001,25(1):110-114.
[12] 邹厚远,程积民,周麟,Hongo A.黄土高原草原植被的自然恢复演替及调节.水土保持研究,1998,5(1):127-138.
[13] 李瑞,张克斌,刘云芳,王百田,杨晓晖,侯瑞萍.西北半干旱区湿地生态系统植物群落空间分布特征研究.北京林业大学学报,2008,30(1):6-13.
[14] 任继周.草业科学研究方法.北京:中国农业出版社,1998:1-30.
[21] 王仁忠.干扰对草地生态系统生物多样性的影响.东北师范大学学报:自然科学版,1996,(3):112-116.
[22] 贾晓妮,程积民,万惠娥.封育对云雾山本氏针茅草地群落的影响.草地学报,2008,16(3):272-277.
[23] 陈芙蓉,程积民,于鲁宁,李媛,吴艳芹.封育和放牧对黄土高原典型草原生物量的影响.草业科学,2011,28(6):1079-1084.
[24] 王炜,刘钟龄,郝敦元,梁存柱.内蒙古草原退化群落恢复演替的研究——Ⅰ.退化草原的基本特征与恢复演替动力.植物生态学报,1996,20(5):449-459.
[25] 祁彪,张德罡,丁玲玲,张力,汪玺,杨予海.退化高寒干旱草地植物群落多样性特征.甘肃农业大学学报,2005,40(5):626-631.
[26] 范永刚,胡玉昆,李凯辉,于建梅,王鑫.不同干扰对高寒草原群落物种多样性和生物量的影响.干旱区研究,2008,25(4):531-536.
[28] 裴成芳,焦金寿.草原火对克氏针茅和扁穗冰草的影响试验.中国草地学报,1998,(3):39-40,46-46.
[31] 李晓波,周道玮,姜世成.火因子对松嫩羊草草原植物多样性的影响.草业科学,1997,14(4):61-64.
[32] 贺郝钰,苏洁琼,黄磊,贾荣亮,李新荣.火因子对荒漠化草原草本层片植物群落组成的影响.生态学报,2011,31(2):364-370.
[33] 周道玮,姜世成,胡勇军.草原植物高生长、体内水分和叶绿素含量对火烧的反应.东北师范大学学报:自然科学版,1999,(4):91-95.
[34] 王辉,谢永生,杨亚利,揣峻峰.云雾山铁杆蒿茎叶浸提液对封育草地四种优势植物的化感效应.生态学报,2011,31(20):6013-6021.
[37] 李伟.施肥导致高寒草甸物种多样性丧失机制研究[D].兰州:兰州大学,2011.
猜你喜欢
天津农林科技(2022年2期)2022-04-19 10:48:14
河南畜牧兽医(2022年3期)2022-04-13 09:12:58
现代园艺(2021年23期)2021-12-01 07:47:44
绿色科技(2021年10期)2021-06-23 03:13:06
林业勘查设计(2020年1期)2021-01-18 02:40:48
新农业(2020年18期)2021-01-07 02:17:08
南方周末(2019-06-27)2019-06-27 19:47:54
草业学报(2019年2期)2019-02-25 01:57:16
小学生作文(低年级适用)(2018年12期)2018-03-23 02:26:14
莫愁(2017年15期)2017-05-11 01:21:55
