
侵染加工番茄的BBWV2的分子鉴定及田间检测
2013-11-02 14:16:34张建云许文博刘升学黄家风
石河子大学学报(自然科学版) 2013年6期
张建云,许文博,刘升学,黄家风
(1石河子大学农学院/新疆绿洲农业病虫害治理与植保资源利用自治区普通高校重点实验室,石河子832003;2大连市农业科学研究院,大连116036)
蚕豆病毒属(Fabavirus)病毒由多种蚜虫以非持久性方式传播,具有广泛的寄主范围,对许多经济重要的农作物及园艺植物造成严重危害[1]。病毒粒体球形,由大、小2种衣壳蛋白组成;基因组由两条正单链 RNA 分子-RNA1(6.0kb)和 RNA2(3.6 kb)组成,采用多聚蛋白翻译策略。由RNA1编码的1个多聚蛋白被进一步加工形成5个功能蛋白,即辅助因子(Co-Pro)、解旋酶(Hel)、蛋白酶(Pro)、基因连锁蛋白(VPg)和依赖于RNA的RNA聚合酶(Pol)。由RNA2编码的1个多聚蛋白被进一步加工形成3个功能蛋白,即运动蛋白(MP)和2个衣壳蛋白(LCP和SCP)[2]。
目前认为,该属有4个种,即蚕豆萎蔫病毒1(Broad bean wilt virus 1,BBWV1)、蚕豆萎蔫病毒2(Broad bean wilt virus 2,BBWV2)、野芝麻轻花叶病毒(Lamium mild mosaic virus,LMMV)和龙胆花叶病毒(Gentian mosaic virus,GeMV)[3]。而曾经报导的广藿香轻花叶病毒(Patchouli mild mosaic virus,Pat-MMV)和薇甘菊花叶病毒(mikania micrantha mosaic virus,MMMV)在第9次国际病毒分类委员会的报告中将其分别列为BBWV2的1个株系和GeMV的1个分离物[2]。与Comoviridae科内豇豆花叶病毒属(Comovirus)和蠕传病毒属(Nepovirus)相比,Fabaviruses分子生物学的研究起步很晚,直到1998年末,日本学者才报道了BBWV2日本分离物的基因组全序列[4]。国内戚益军等[5]于2000年首次获得BBWV2中国分离物的基因组全序列,目前已有越来越多的BBWV1和BBWV2被克隆和测序。BBWV1和BBWV2除了基因组序列有差异,其抗原表位分属不同的血清型[6]。从发生分布来看,BBWV1主要分布于欧洲、美洲等,对辣椒、蚕豆、菠菜等造成危害。BBWV2主要分布于东亚,侵染辣椒、蚕豆及一些中草药[1,7],在我国迄今尚未分离到BBWV1。
加工番茄是新疆重要的经济作物,但是病毒病的发生与危害一直是影响其产量的重要因素。随着农产品贸易的增强、耕作制度的改变和传毒介体的多样化,田间病毒症状及发生危害日趋复杂,已有多种病毒被鉴定出来,如黄瓜花叶病毒(Cucumber mosaic virus,CMV)、番茄花叶病毒(Tomato mosaic virus,ToMV)、马铃薯 Y 病毒(Potato virus Y,PVY)、马铃薯 M 病毒(Potato virus M,PVM)、中国番茄黄化曲叶病毒(Tomato yellow leaf curl Chi-na virus,TYLCCNV),其中CMV和ToMV通常以复合侵染的方式在田间造成危害[8-11]。
本研究通过RT-PCR、克隆和测序对田间病株上发生的BBWV2进行了分子鉴定,并通过ELISA对田间病株的发生情况进行了调查,旨在为该病害防治提供依据。
1 材料与方法
1.1 材料
1.1.1病毒来源
用于病毒种类分子鉴定的23个田间病株于2009年采自石河子大学农学院试验站晚播加工番茄;用于ELISA检测的田间病株于2010-2011年采自石河子大学农学院试验站和石河子蔬菜所。
1.1.2供试抗血清
BBWV2的单克隆抗体由浙江大学生物技术研究所周雪平教授惠赠。
辣根过氧化物酶标记山羊抗小鼠IgG购自北京博奥森生物技术有限公司。
酶标板(costar 3590)购自美国Corning公司,酶标仪为日本BIO-RAD Model 550型。
1.2 方法
1.2.1总RNA提取
参照改良的LiCl方法[12]提取加工番茄病株总RNA。
1.2.2 RT-PCR扩增、克隆和序列测定
1)以病株的总RNA作为模板,用蚕豆病毒属(Fabavirus)的兼并引物 Fab5′R1F和Fab5′R1R[13]对病株总RNA进行RT-PCR,选取代表病株PCR产物进行克隆、测序。
2)根据已测序列及GenBank中登录的BBWV2的RNA1序列保守区设计了3对引物对XJ14-1基因组RNA1进行分段RT-PCR。
3)根据基因库中已登陆BBWV2的RNA2核苷酸序列设计了1对引物R2F-1F和R2F-1R,对基因组RNA2部分序列进行RT-PCR。
4)将PCR产物分别克隆到载体pMD18-T vector(TaKaRa,Japan)上,选择PCR和酶切鉴定均为阳性的克隆。
委托上海生工生物工程技术服务有限公司测序。所用引物序列见表1。

表1 用于扩增BBWV2基因组RNA1和RNA2的引物Tab.1 Primers used for amplifying of RNA1and RNA2of BBWV2
1.2.3序列分析
测序结果提交NCBI进行BLAST检索。
利用软件DNAStar 5.02和DNAMAN Version 5.2.2进行序列分析。
多序列比较采用DNAStar clustal W方法,系统进化树使用 MEGA5.0(Amazon Inc.Masatoshi Nei and Sudhir Kumar,USA)程序进行构建。
1.2.4抗原制备
将田间采集的病株叶片在研钵中磨碎,用0.1 mol/L磷酸缓冲液 (pH 7.5)按1∶10的比例稀释,4000r/min离心5min,吸取上清液备用;同时以加工番茄健康叶片提取液作阴性对照。
1.2.5 ELISA检测
采用间接ELISA方法,用稀释的待测抗原包被微量反应板16-18h,封闭后,加适量稀释的已知单克隆抗体,反应后再加适当浓度的酶标抗体,经TMB底物反应显色,加2mol/L硫酸终止。然后在BIORAD Model 550型酶标仪上,于490nm处,以空白对照孔调零后测各孔OD值,以P/N>2.1作为阳性判断标准(P为待测样品OD值,N为阴性对照OD值)。每个样品设3次重复。
2 结果与分析
2.1 田间病株BBWV2的RT-PCR检测
用蚕豆病毒属(Fabavirus)的兼并引物Fab5′R1F和Fab5′R1R对23个病株进行RT-PCR扩增,结果显示,19个病株扩增到预期大小的约400bp的DNA片段(图1)。
选取其中 XJ7-5、XJ7-6、XJ7-9和 XJ14-1的PCR产物分别回收、克隆和序列测定,得到目标片段的核苷酸(nucleotide,nt)序列分别为391、390、392和390 nt;4个序列之间的同源性为95.1%~98.2%。
基因库(http://www.ncbi.nlm.nih.gov/)检索结果表明,4个序列均与蚕豆萎蔫病毒2(BBWV-2)基因组RNA1组分5′末端第11位核苷酸开始的390~392 nt序列同源性最高,说明经RT-PCR检测,具有大小约400bp片段的田间病株均被BBWV2侵染。
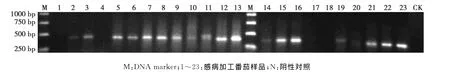
图1 感病加工番茄样品蚕豆病毒属病毒的RT-PCR检测Fig.1 RT-PCR amplification of Fabavirus from processing tomato samples showing viral symptom
2.2 病毒分离物XJ14-1基因组RNA1和RNA2的测序及同源性比较
为了更多了解侵染加工番茄的BBWV2的基因组信息,根据上述检测结果,选取病株XJ14-1,对其病毒分离物基因组RNA1和RNA2进行克隆与测序。首先对基因库中已登陆BBWV2的RNA1核苷酸序列进行比对,设计了3对引物对基因组RNA1进行分段扩增(表1),结果如图2所示。
图2显示:利用R1F-1F和R1F-1R引物扩增到约1.2kb的特异性片断;用 R1F-31F和 R1F-31R引物扩增到约1.5kb的特异性片断;用R1F-32F和R1F-32R引物扩增到约1.7kb的特异性片断。
将各目标片段克隆、测序,得到核苷酸序列分别为1185、1517、765nt(该片段部分序列未被测出),再将这3个序列与之前测得的390nt拼接,获得基因组RNA1组分5′末端第11位核苷酸开始的的3659个核苷酸。再将该序列与基因库中已登陆的BBWV2的RNA1进行序列比对,结果见表2。
由表2可知:它与韩国RP3分离物的同源性最高,达到94%,与其它BBWV2的同源性在76.6%~89.7%,与BBWV1的RNA1的同源性仅为54.1%。
根据基因库中已登陆BBWV2的RNA2核苷酸序列设计了1对引物R2F-1F和R2F-1R对基因组RNA2部分序列进行了RT-PCR、克隆和测序,获得1300nts的核苷酸序列结果见图3。将该序列与基因库中已登陆的BBWV2的RNA2进行序列比对,结果见表2。
由表2可知:该序列与韩国K分离物的同源性最高,达到94%,而与韩国RP3分离物的同源性为83.4%,与其它BBWV2的同源性为76.6%~86.3%,与BBWV1的RNA2的同源性很低,仅为48%。

表2 病毒分离物XJ14-1与其它蚕豆萎蔫病毒核苷酸序列相似性比较Tab.2 Comparison of nucleotide sequence identities of RNA1and RNA2between XJ14-1and other Fabavirus
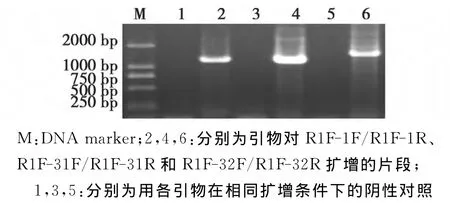
图2 XJ14-1基因组RNA1的分段扩增Fig.2 Amplification of the XJ14-1genome segment RNA1
2.3 田间病株BBWV2的ELISA检测
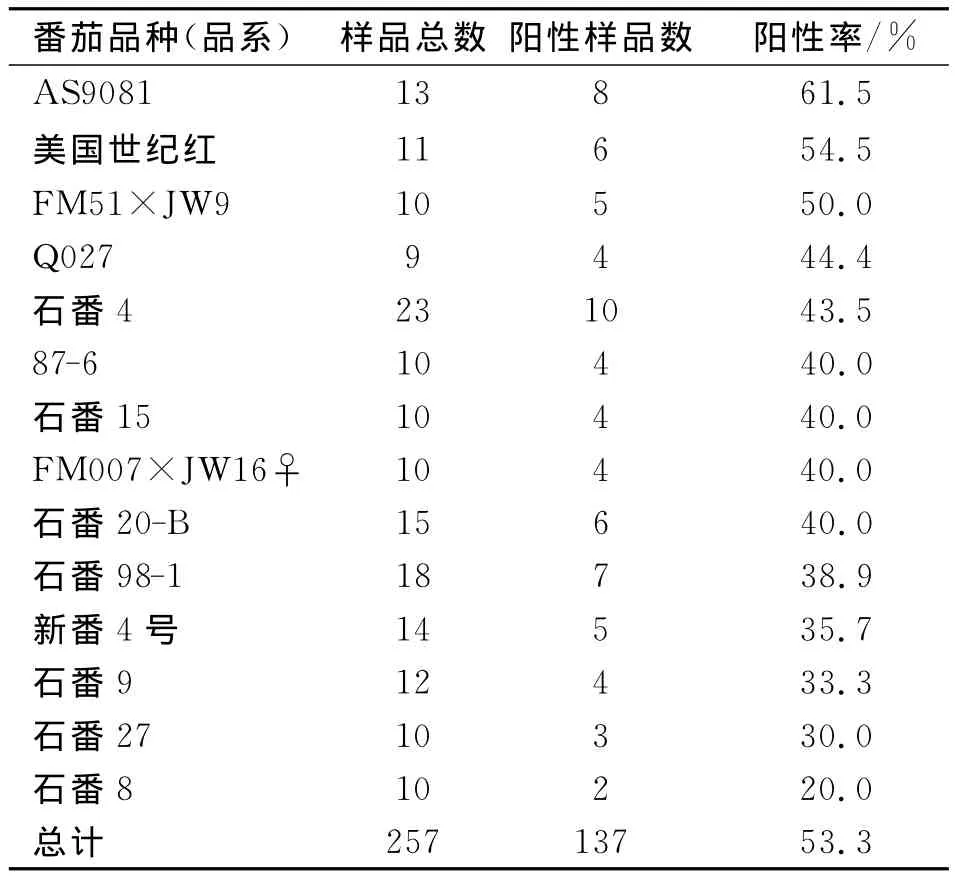
在明确加工番茄可被BBWV2侵染的基础上,利用BBWV2的单克隆抗体对田间采集的不同品种(品系)加工番茄病株进行了ELISA检测,结果见表3。
表3显示,20个品种都能检测到BBWV2,阳性率为20%~100%。257个供试病株,有137个病株检测到BBWV2,总阳性率为53.3%。由此说明,田间病株带毒率达到53.3%,并且供试20品种(品系)都可被BBWV2侵染。
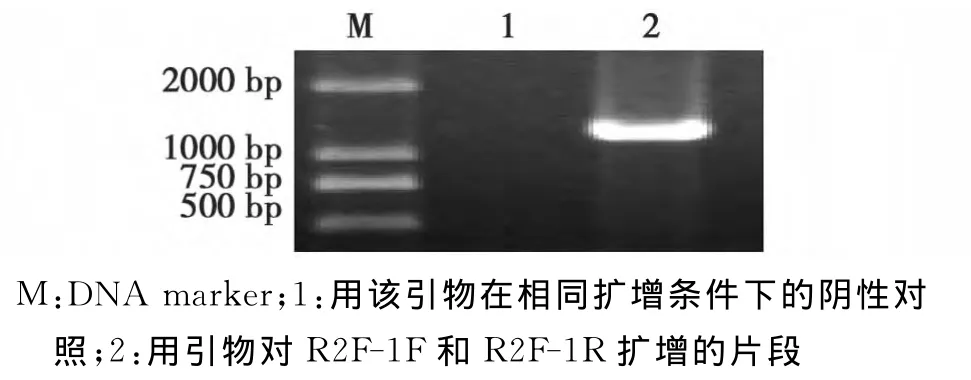
图3 XJ14-1基因组RNA2的分段扩增Fig.3 Amplification of the XJ14-1genome segment RNA2
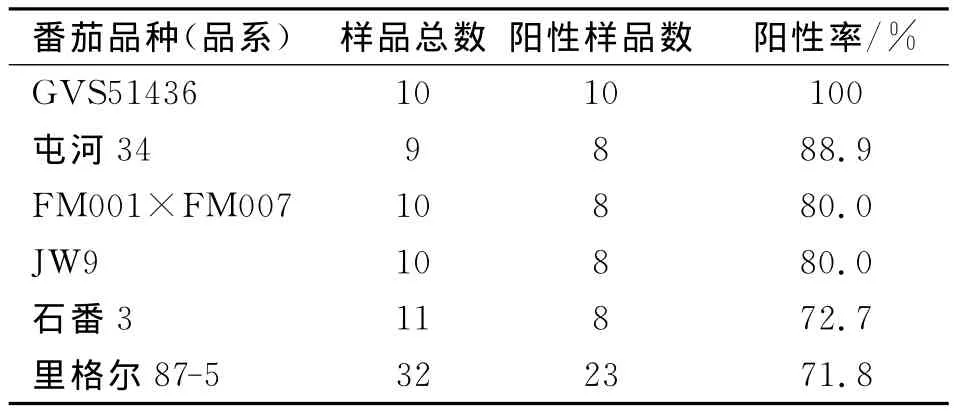
表3 田间病株BBWV2的ELISA检测结果Tab.3 Detection of field samples showing viral symptom by ELISA
续表3
3 结论与讨论
1)有关蚕豆萎蔫病毒在新疆的最早危害始于1994年,由周俊等研究[14],他们从表现斑驳的番茄病株上分离到1种球状病毒,通过对病毒的症状特征、寄主范围、衣壳蛋白氨基酸组成等初步判断该病毒为蚕豆萎蔫病毒(BBWV),却不能明确病毒种类是BBWV1还是BBWV2。蚕豆病毒属(Fabavirus)的兼并引物既能作为检测引物从病株上检测该属病毒,又能通过RT-PCR扩增的片段大小有效区分该属3种病毒(BBWV1、BBWV2和 GeMV)[13]。
本研究利用该对引物通过RT-PCR从23个加工番茄病株上检测出19株被病毒侵染,然后通过对其中1个病毒分离物XJ14-1的基因组RNA1和RNA2进行测序及序列比对,结果表明XJ14-1的基因组与GenBank中登录的BBWV2都具有高的序列同源性,而与BBWV1的同源性却很低,因此从分子水平明确新疆加工番茄受到BBWV2的侵染。
2)利用BBWV2的单克隆抗体对田间不同品种(品系)加工番茄病株进行ELISA检测,结果显示:田间病株带毒率达53.3%,且供试20品种(品系)都可被BBWV2侵染;所得数据表明,BBWV2在田间发生普遍,是危害新疆加工番茄的主要病毒之一。
由于BBWV2寄主分布很广,我国已先后在多种豆科植物及百脉草、紫云英、辣椒等多种植物上发现有该病毒侵染[15]。这些寄主有的是BBWV2的越冬寄主,有的是桥梁寄主,它们为病毒周而复始的循环危害提供了场所及栖息地。因此,对该病毒在新疆的发生情况还需进行更广泛的调查分析。
[1]Ferrer R M,Ferriol I,Moreno P,et al.Genetic variation and evolutionary analysis of broad bean wilt virus 2[J].Archives Virology,2011,156(8):1445-1450.
[2]Sanfacon H,Iwanami T,Karasev A V,et al.Virus taxonomy,ninth report of the international committee on the taxonomy of viruses[M].London:Elsevier Academic Press,2011,881-899.
[3]Dong S W,Xiang H Y,Shang Q X,et al.Complete genomic sequence analysis reveals a novel fabavirus infecting cucurbits in China[J].Archives Virology,2012,157(3):597-600.
[4]Nakamura S,Iwai T,Honkura R.Complete nucleotide sequence and genomic organization of Broad bean wilt virus 2[J].Ann Phytopathol Soc Jpn,1998,64:565-568.
[5]Qi Y J,Zhou X P,Li D B.Complete Nucleotide Sequence and Infectious cDNA Clone of the RNA1of a Chinese I-solate of Broad Bean Wilt Virus 2[J].Virus Genes,2000,20(3):201-207.
[6]Murphy F A,Fauquet C M,Bishop D H L,et al.Virus Taxonomy:Classification and Nomenclature of Viruses.Sixth report of the International Committee on Taxonomy of Viruses[M].Vienna:Springer-Verlag,1995,341-347.
[7]Ferriol I,Ruiz-Ruiz S,Rubio L.Detection and absolute quantitation of Broad bean wilt virus 1(BBWV-1)and BBWV-2by real time RT-PCR [J].Journal of Virological Methods,2011,177(2):202-205.
[8]许文博,都业娟,黄家风.加工番茄CMV与ToMV的ELISA检测及其相关性分析[J].石河子大学学报:自然科学版,2009,27(2):199-201.
[9]姜玉霞,向本春,安仙丽.新疆加工番茄上番茄花叶病毒的分子鉴定[J].新疆农业科学,2008,45(3):484-489.
[10]都业娟,许文博,向本春,等.侵染新疆加工番茄的中国番茄黄化曲叶病毒DNA-A的基因组特征[J].植物病理学报,2011,41(4):393-398.
[11]崔燕华,向本春.新疆加工番茄顶端叶片黄化现象的初步研究[J].石河子大学学报:自然科学版.2011,29(4):425-431.
[12]常波,向本春,刘升学,等.加工番茄条斑坏死病原黄瓜花叶病毒外壳蛋白基因的克隆和序列分析[J].石河子大学学报:自然科学版,2006,24(4):410-414.
[13]Ferrer R M,Luis-Arteaga M,Guerri J,et al.Detection and identification of species of the genus Fabavirus by RT-PCR with a single pair of primers[J].Journal of Virological Methods,2007,144(1-2):156-160.
[14]周俊,尹玉琦,崔星明,等.蚕豆萎蔫病毒-新疆番茄分离株的鉴定[J].中国病毒学,1994,9(4):327-331.
[15]周雪平,余永杰,戚益军.蚕豆萎蔫病毒ELISA检测系统建立及其应用[J].植物病理学报,1996,26(4):347-352.
猜你喜欢
当代水产(2022年1期)2022-04-26 14:35:30
小读者(2021年2期)2021-03-29 05:03:30
新农民(2020年22期)2020-08-31 01:32:08
公民与法治(2020年13期)2020-07-27 01:30:36
科学咨询(2020年10期)2020-04-01 03:43:16
乡村科技(2019年25期)2019-02-21 07:40:48
学生天地(2018年9期)2018-09-06 13:11:18
现代园艺(2017年11期)2017-06-28 11:32:37
西南农业学报(2016年5期)2016-05-17 05:42:33
西南农业学报(2016年6期)2016-04-16 05:12:51
