
不同围封年限克氏针茅群落物种多样性分析
2013-10-31 02:26:30白文科王志军张昊巴布拉
草原与草业 2013年3期
白文科,王志军,2,张昊* ,巴布拉
1 引言
群落物种多样性是群落的重要特征,任何一种干扰因子对群落结构的影响都离不开物种多样性问题。克氏针茅(Stipakrylovii)为多年生密丛型旱生草本植物,是亚洲中部草原典型植被的主要建群种〔1〕。克氏针茅草原是典型草原带和荒漠区山地草原带的地带性群系,也是某些大针茅草原的放牧演替变型〔2〕。克氏针茅草原,是内蒙古高原典型草原地带的优势群系,是天然放牧场主要的草原植被类型〔3〕。随围封年限的延长,群落盖度与密度增加〔4〕。因此不同围封年限克氏针茅群落物种多样性的分析对退化草地的恢复及恢复过程中的合理利用,草地资源的合理利用与有效管理、生物多样性的保护及草地畜牧业的可持续发展等等方面都具有较为重要的意义。
为了使已经退化的草原生态系统逐步得到恢复,保护草原丰富的物种多样性,人们采取了各种各样的保护措施〔5〕,其中对退化草地实行围栏禁牧恢复是区域生态重建的一项重要举措〔6〕。草地围栏封育由于其投资少、见效快,已成为当前退化草地恢复与重建的重要措施之一〔7〕。但是,到目前为止,有关禁牧对草原生态系统影响的研究相对来说还比较少见,尤其是有关禁牧与草原生态系统物种多样性关系的研究,而生态系统中生物多样性的维持是生态系统管理的中心目标之一〔8〕。
2 研究地区与研究方法
2.1 研究区概况
研究地区为内蒙古锡林郭勒盟正镶白旗,地理坐标为东经 114°05'-115°37',北纬 42°05'-43°15'、海拔高度一般在1200-1400m之间;年平均温度1.9℃;年均降水量300mm左右;年均蒸发量2000mm;无霜期125d;日照时数3044h;;年平均风速4m/s,全年大风日数78d(6-8级);土壤以栗钙土为主。植被类型由于受气候、土壤基质的影响,特别是水热分配的差异,形成了以地带性干草原为主体,隐域性沙地、草甸植被次之的植被类型。
2.2 研究方法
2.2.1 野外调查
通过对克氏针茅草原踏查后,在不同退化地段上选取标准样地,做1×1m2样方,做描述样方和记名样方,描述样方5次重复,记名样方20次重复,分别调查每个样方内的植物种类、高度(H)、密度(D),频度(F)、盖度(C)和产量(P)等,7月初开始进行群落结构调查,获取群落物种多样性资料。
(1)样线法:在每个试验样地内取2条平行的100m的直线,沿线记录此线所遇到的植物,并分析。
(2)样方法:沿6条100m的样线,每条线设置5个1m×1m的观测样方。采用目测法确定群落总盖度,用直尺测量每种植物的平均高度。将其地上绿色部分齐地面刈割并分种存放于塑封袋中,将其立枯与凋落物分别收集于塑封袋中。带回实验室分别称量其鲜重。然后风干称量其干重。
2.2.2 内业整理
在内蒙古农业大学生态环境学院草地实验室内对采集的样线、样方数据及地上生物量进行称量和总结;并在内蒙古农业大学生态环境学院资料室内对《典型草原植物多样性研究》、《内蒙古自治区植物志》、《生态学报》等相关书籍及相关文献进行查阅,使得本次研究更加准确。
(1)群落重要值的计算方法
物种重要值计算公式如下:IV=RDE+PHI+PCO,
其中,IV为重要值;RDE为相对密度;RHI为相对高度;RCO为相对盖度
(2)群落物种多样性的计算方法
计算公式如下:
(1)丰富度指数:采用Margalef丰富度指数:R=(S-1)/ln
(2)多样性指数:采用Shannon-Winner多样性信息指数:H=-∑Piln(Pi)
(3)优势度指数:采用Simpson优势度指数:C=1-∑(Pi)2
(4)均匀度指数:采用Pielou均匀度指数:E=-∑Piln(Pi)/ln(S)
其中,S为群落物种数目,N为所有物种个体数,Pi:IV/IV∑。
3 结果与分析
3.1 不同围封年限克氏针茅群落物种变化情况
经过样线调查,在围封五年的草场其物种组成主要有糙隐子草、短花针茅、克氏针茅、羊草;围封十年(以上)主要植物种有阿氏旋花、并头黄芩、糙隐子草、短花针茅、克氏针茅、冷蒿、羊草。
在围封五年草场平行的两条样线上有:糙隐子草52株,平均高度为4.78cm、短花针茅317株,平均高度为 8.29cm、克氏针茅 83株,平均高度为9.70cm、羊草39株,平均高度为8.49cm。围封十年(以上)草场平行的两条样线上有:阿氏旋花82株,平均高度为7.71cm,并头黄芩78株,平均高度为9.09cm,糙隐子草76株,平均高度为9.07cm、短花针茅23株,平均高度为14.49cm、克氏针茅343株,平均高度为28.62cm、冷蒿 58株,平均高度为13.12cm,羊草71株,平均高度为19.41cm。(图1)。
不同围封年限草地,主要植物群落植物种组成,密度及高度均呈现不同的变化特征。从群落植物种组成,密度及高度看,不同围封年限后,群落植物的密度、高度、地上部分生物量与主要物种组成均表现为有所增加。
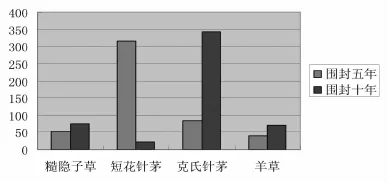
图1 不同围封年限样线植物数对比图
因此,在退化草地恢复过程中,群落植物高度、密度、生物量、群落物种组成成分弹性均较大。虽然不同围封年限群落各项指标显著性地增加,但克氏针茅和短花针茅密度及生物量增长幅度并不是呈现等比例增加,这说明不同植物生物量恢复增长弹性与围封年限不是成比例增长,这可能是经过长期恢复演替该区植物群落已接近某种顶极状态,因而个别植物生物量增长不是等比例增长,甚至趋于减少状态。
3.2 克氏针茅群落中主要物种重要值的变化情况
群落物种重要值可用来反映其在群落中的优势地位及优势程度,故可作为表征优势种的指标。本研究中的数据是在同一区域不同围封年限克氏针茅草原调查统计的。
不同围封年限克氏针茅群落主要物种重要值如表1所示:
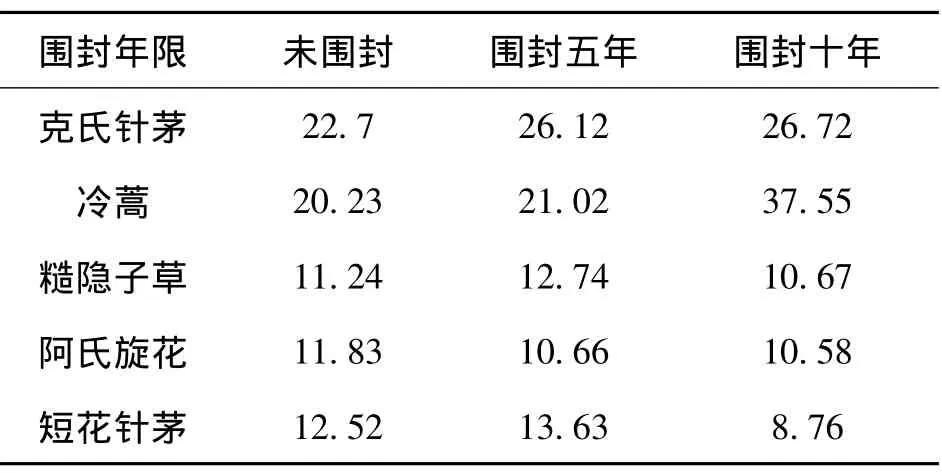
表1 群落中主要物种重要值(%)
据表1可看出,在不同围封年限克氏针茅群落中,克氏针茅,冷蒿等占有较大优势,但随着围封年的减少,阿氏旋花的重要值增大,在未围封地区达最大值;短花针茅则在围封十年地区重要值明显降低;糙隐子草的重要值先增后减,在五年围封地区达最大值;而克氏针茅和冷蒿的重要值在十年围封区达最大值。
3.3 不同围封年限克氏针茅群落多样性指数的变化情况
植物群落多样性一般指植物群落在组成、结构、功能和动态方面所表现出的差异。a多样性是物种多样性,是指物种种类与数量的丰富程度,是一个区域或一个生态系统可测定的生物学特征。不同围封年限克氏针茅群落a多样性指数如表2所示:
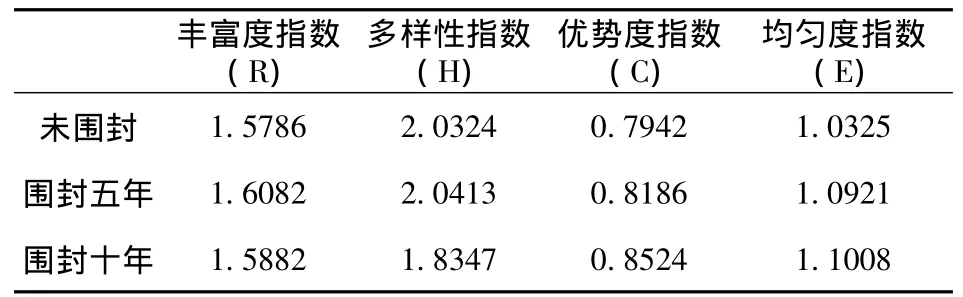
表2 不同围封年限克氏针茅群落物种多样性指数
据表2可以看出,丰富度指数,优势度指数,表现为围封五年区最高,同时优势度指数和均匀度指数十年围封区最高。
3.4 不同围封年限对克氏针茅数量特征的影响
不同围封年限对克氏针茅数量特征的影响随着围封年限的延长,克氏针茅种群中个体的数量特征呈先增加后减小的趋势。在围封5-10年间,克氏针茅种群的个体数量特征:如生殖枝高度、营养枝长度、生殖枝干重等都比未围封样地克氏针茅个体的数量特征有显著增加;随着围封年限延长而这些指标不变或者显著减小,在围封十年样地克氏针茅种群的个体数量特征:如营养枝长度、生殖枝干重等指标显著小于围封5-10年样地克氏针茅个体的指标。生殖枝密度和生殖枝/总生物量随着围封时间的延长呈先增加后降低的趋势,生殖枝密度和生殖枝干重/总生物量在围封五年样地中显著高于其他样地。克氏针茅种群密度和单位面积生物量随着围封年限的延长呈先增加然后逐渐降低的趋势,克氏针茅种群密度和单位面积生物量在围封五年样地中显著高于其他样地。
4 结论
(1)不同围封年限克氏针茅群落物种重要值表现为:克氏针茅和冷蒿在十年围封区达最大值;糙隐子草在五年围封区达最大值;阿氏旋花在未围封地区达最大值。
(2)围封5年的克氏针茅群落,物种的丰富度、均匀度和多样性均增加。随围封年限的延长,物种丰富度继续增加,但由于围封10年后形成单优势种群落,群落多样性降低。之后,随围封年限的继续增加,群落逐步趋于稳定,丰富度略为降低,均匀度和多样性增加。另外,从群落相似性来看,采用生长季围封恢复措施后,围封样地与未围封对照间的相似性降低,说明季节性围封不仅能增加群落物种多样性,而且在改变群落的环境条件上也具有较好的效果。
(3)在本研究中,不同围封年限的克氏针茅群落,其植物群落的物种组成、结构和生物多样性特征发生了不同程度的改变。未围封样地,其群落结构简单,盖度小,群落的多样性、丰富度和均匀度逐步降低,而群落优势度逐渐增大。虽然各退化草地都形成了相对稳定的群落结构,但优势物种和植物群落多样性的变化却改变了草原生产力的可利用性。另一方面,对本区研究表明对草地实行围栏封育,禁止放牧利用,虽有利于群落优势度指数增大,但系统内群落的物种丰富度、均匀度和植物多样性均显著低于适度利用的草地,而适度放牧利用草地的群落物种多样性和丰富度以及植被均匀度均表现为最高。
〔1〕祁彪,张德罡,丁玲玲,张力,汪玺,杨予海.退化高寒干旱草地植物群落多样性特征〔J〕.甘肃农业大学学报,2005,10:626-631.
〔2〕刘珏宏,高慧,张丽红,陈丽萍,赵念席,高玉葆.内蒙古锡林郭勒草原大针茅-克氏针茅群落的种间关联特征分析〔J〕.植物生态学报,2010,34(9):1016-1024.
〔3〕吴建波,包晓影,李洁,赵念席,高玉葆.不同围封年限对典型草原群落及大针茅种群特征的影响〔J〕.草地学报,2010,7.
〔4〕刘美珍,蒋高明,于顺利,李永庚,高雷明,牛书丽,姜闯道,彭羽.浑善达克退化沙地恢复演替18年中植物群落动态变化〔J〕.生态学报,2004,8.
〔5〕韩冰,赵萌莉,珊丹.不同退化梯度克氏针茅种群形态及等位酶的分析〔J〕.草业科学,2004,12:78.
〔6〕许中旗,李文华,许晴,闵庆文,王英舜,何旭生.禁牧对锡林郭勒典型草原物种多样性的影响〔M〕.生态学杂志,2008,27(8):1307-1312.
〔7〕周华坤,周立,刘伟,王启基,赵伟,周彦艳.封育措施对退化与未退化矮蒿草草甸的影响〔J〕.中国草地,2003,9.
〔8〕单贵莲,徐柱,宁发,马玉宝,李临杭.围封年限对典型草原群落结构及物种多样性的影响〔J〕.草业学报,2008,12:1-8.
猜你喜欢
作物研究(2023年2期)2023-05-28 13:44:14
林业勘查设计(2022年1期)2022-02-15 05:35:12
乡村科技(2021年17期)2021-10-20 08:50:56
安顺学院学报(2021年4期)2021-09-16 08:11:08
林业调查规划(2020年3期)2020-06-03 07:02:36
河南畜牧兽医(2020年21期)2020-01-10 00:20:08
纺织科技进展(2015年1期)2015-11-28 05:56:28
实用手外科杂志(2015年4期)2015-08-27 01:54:08
实用手外科杂志(2015年3期)2015-08-27 01:53:14
实用手外科杂志(2015年3期)2015-08-27 01:53:14
