
藻胆蛋白的提取、纯化及其体外抗紫外活性
2013-10-16 01:48叶翠芳张美英汤顺清王一飞利奕成
暨南大学学报(自然科学与医学版) 2013年5期
叶翠芳, 杨 珂, 张美英, 汤顺清, 王一飞, 利奕成
(暨南大学1.生物医药研究开发基地;2.生物医学工程系,广东广州510632)
紫菜,属红藻门,原红藻纲,红毛菜目,红毛菜科,紫菜属海藻植物的统称,含有高蛋白质、多种维生素、碳水化合物等营养成分.藻胆蛋白是一种具有多种生物活性的水溶性色素蛋白,广泛存在于海洋藻类植物中.藻胆蛋白色泽鲜艳、颜色特殊,具有避免化学合成物质对人们身体伤害的作用,欧美等发达国家很早就把藻胆蛋白用作了食品的色素添加剂或者化妆品的生产[1],除此之外,藻胆蛋白还具有增强免疫力、抗氧化、抗衰老等多种生物活性.
UVA与皮肤光老化关系密切,而成纤维细胞是紫外线引发皮肤光老化的主要作用靶点,紫外线对成纤维细胞的主要表现在直接与生物大分子发生反应,引起DNA的损伤,从而造成成纤维细胞凋亡,数量减少,活性降低而导致胶原合成减少,致使皮肤张力和承受拉力降低[2].藻类是世界公认的抗辐射物质,目前对于海藻中天然紫外线吸收物质的研究还处于起步阶段,大多数研究主要集中在环境条件对藻类积累紫外吸收物质的影响及紫外线对其的诱导等方面[3].
我国藻类资源丰富,是一个紫菜生产大国,是获取藻胆蛋白的理想资源,选用紫菜提取藻胆蛋白具有成本低、得率高的优点,具有较好的利用价值.由于藻胆蛋白具有很好的社会应用前景,本研究拟从紫菜中提取分离藻胆蛋白,建立体外抗紫外损伤细胞实验模型,从细胞学水平评价藻胆蛋白的抗紫外效果,为其在紫外防护、细胞修复等方面的应用提供依据.
1 实验部分
1.1 材料
干紫菜(产地福建,第一水品质);小鼠成纤维细胞(NIH 3T3,武汉大学保藏中心);96孔微量细胞培养板(美国corning公司);胎牛血清(FBS,杭州四季清公司);青霉素钠、硫酸链霉素(华北制药股份有限公司);四甲基偶氮唑盐、二甲基亚砜(分析纯,美国SIGMA公司);硫酸铵、盐酸、磷酸氢二钠、磷酸二氢钾、氯化钠、氯化钾(分析纯,广州化学试剂厂).
1.2 仪器
冷冻离心机(德国Hettich);紫外可见分光光度计(上海美普达仪器有限公司);层析柱(上海锦华层析设备厂);核酸蛋白检测仪(上海嘉鹏科技有限公司);ECP3000三恒电泳仪(北京六一仪器厂);UVA灯管(PHILIPS公司);二氧化碳培养箱(日本SANYO);显微镜(日本OLYMPUS);超净工作台(苏州苏净集团);酶标自动读数仪(美国BIO-RAD);流式细胞分析仪(BD Calibur).
1.3 实验方法
(1)藻胆蛋白的提取[4-7]称取干紫菜15 g,用1 L去离子水浸泡15 min使之溶胀,在冰浴中进行超声波破碎,然后先用100目滤布过滤,再将滤液在4℃下离心(9 000 r/min,10 min),去除滤渣得到紫红色的上清液.
量取上清液体积,先用质量分数25%饱和硫酸铵沉淀杂蛋白,4℃静置2 h后离心收集沉淀并用适量的水复溶,然后用质量分数60%饱和硫酸铵继续沉淀,4℃静置2 h后离心收集沉淀,将沉淀用PBS溶解并脱盐处理,得到藻胆蛋白粗提液.
(2)藻胆蛋白的纯化[8-10]把DEAE-纤维素填料装入层析柱,用20 mmol/L磷酸钠缓冲液(pH=6.5)平衡,上样,用含不同浓度NaCl的磷酸钠缓冲液(20 mmol/L,pH=6.5)洗脱,根据洗脱曲线收集洗脱液,并且测定洗脱液的Amax/A280.选取纯度较高的洗脱液,脱盐,再用羟基磷灰石(HAP)层析柱纯化,用 20 mmol/L,50 mmol/L,100 mmol/L 磷酸钠缓冲液(pH=7.0,0.1 mol/L NaCl)洗脱,收集洗脱峰,测Amax/A280,将目标组分脱盐处理并进行紫外光谱扫描和SDS-PAGE分析.
(3)藻胆蛋白对体外培养细胞的影响
①藻胆蛋白对NIH 3T3细胞生长的影响 消化NIH 3T3细胞,以8000/孔接种于96孔培养板中,置于37℃、体积分数5%CO2的细胞培养箱培养24 h后,弃上清,用无血清的DMEM培养基稀释藻胆蛋白,将系列质量浓度(8.65、4.32、2.16、1.08、0.54、0.27、0.14 μg/mL)的藻胆蛋白稀释液分别加入培养孔中,每孔100 μL;同时设正常细胞对照组,加入无血清的DMEM培养基,每孔100 μL.然后置于细胞培养箱(37℃、体积分数5%CO2)中培养.培养48 h 后加入 10 μL/孔的 MTT,继续培养 4 h,弃上清,加入 DMSO 100 μL/孔,避光震荡 15 min,在酶联免疫检测仪上测定其吸收值(检测波长为570 nm,参考波长630 nm),计算细胞存活率以评价藻胆蛋白对NIH 3T3细胞生长的影响.
②NIH 3T3细胞UVA损伤实验造模[11]随机将NIH 3T3细胞分为6组:正常细胞对照组和5个UVA损伤组.探索UVA损伤实验造模的辐射剂量.
小鼠NIH 3T3成纤维细胞培养至融合度70%~80%,各UVA损伤组用UVA紫外线照射细胞,细胞与辐照光源距离15 cm,辐射强度为540 μW/cm2,辐照时间分别为 15、20、30 min、1、2 h,然后继续培养48 h,在显微镜下观察辐照后的各UVA损伤组的细胞形态,根据细胞生长状态,确定NIH 3T3细胞UVA损伤组的造模剂量.
紫外线辐射剂量=辐射强度×辐射时间[12].
③藻胆蛋白体外抗UVA辐射实验 小鼠NIH 3T3成纤维细胞随机分为对照组(Control)、UVA损伤组、藻胆蛋白组、UVA损伤+藻胆蛋白组.UVA损伤组要按②造模实验结果,接受造模剂量的UVA照射.UVA损伤+藻胆蛋白组为UVA损伤组按照实验设计添加系列质量浓度(8.65、4.32、2.16、1.08、0.54、0.27、0.14 μg/mL)的藻胆蛋白稀释液获得,共7个藻胆蛋白剂量;藻胆蛋白组不经UVA照射,按照实验设计添加系列质量浓度的藻胆蛋白稀释液获得,亦分7个小组.
小鼠NIH 3T3成纤维细胞至融合度70%~80%,UVA损伤+藻胆蛋白组细胞待紫外线辐照构建UVA损伤细胞后,弃上清,用无血清的DMEM培养基稀释藻胆蛋白至终质量浓度制备稀释液,加入到构建好的UVA损伤细胞中培养,依据藻胆蛋白的质量浓度,共分为7个小组;藻胆蛋白组不经UVA损伤,其余处理同UVA损伤+藻胆蛋白组,亦分7个小组;对照组、UVA损伤组加入无血清的DMEM培养基.继续培养48 h,加入10 μL/孔的MTT,继续培养4 h,弃上清,加入 DMSO 100 μL/孔,避光震荡15 min,酶标仪读数(波长570 nm,参考波长630 nm),测吸光值,计算细胞存活率.
④流式细胞仪分析细胞周期 实验细胞随机分为UVA损伤组(UVA)、UVA损伤+藻胆蛋白组(UVA+PBP).当小鼠成纤维细胞至融合度80%时,将实验细胞放到UVA紫外灯下照射损伤,弃上清,然后UVA损伤+藻胆蛋白组加入用无血清DMEM培养基稀释至8.65 μg/mL的藻胆蛋白溶液,UVA损伤组则加入无血清DMEM培养基,继续培养48 h后,收集细胞,离心(1 500 r/min,10 min),调整细胞密度为每毫升1×106个细胞,体积分数75%预冷乙醇固定过夜,染色后采用流式细胞仪检测细胞周期,结果用细胞周期拟合软件ModFit LT分析.
2 结果与分析
2.1 藻胆蛋白的纯化结果
(1)藻胆蛋白粗提液经DEAE-纤维素柱纯化,用氯化钠离子梯度洗脱,洗脱曲线见图1.
DEAE-纤维素柱层析纯化得到两个组分,根据各组分的特征吸收及所呈现的颜色辨别,组分1呈蓝紫色,在616、558、652 nm 有吸收峰,主要含有藻蓝蛋白和藻红蛋白,A616/A280和 A558/A280在1.5~2.0;组分2呈亮粉红色,在558、616 nm有吸收峰,主要含有藻红蛋白.收集纯度较高的组分1,再用羟基磷灰石柱层析纯化,洗脱曲线见图2,收集到两个洗脱峰,经测定,洗脱液的纯度A616/A280可达到3.0以上,A558/A280也在2.0以上.
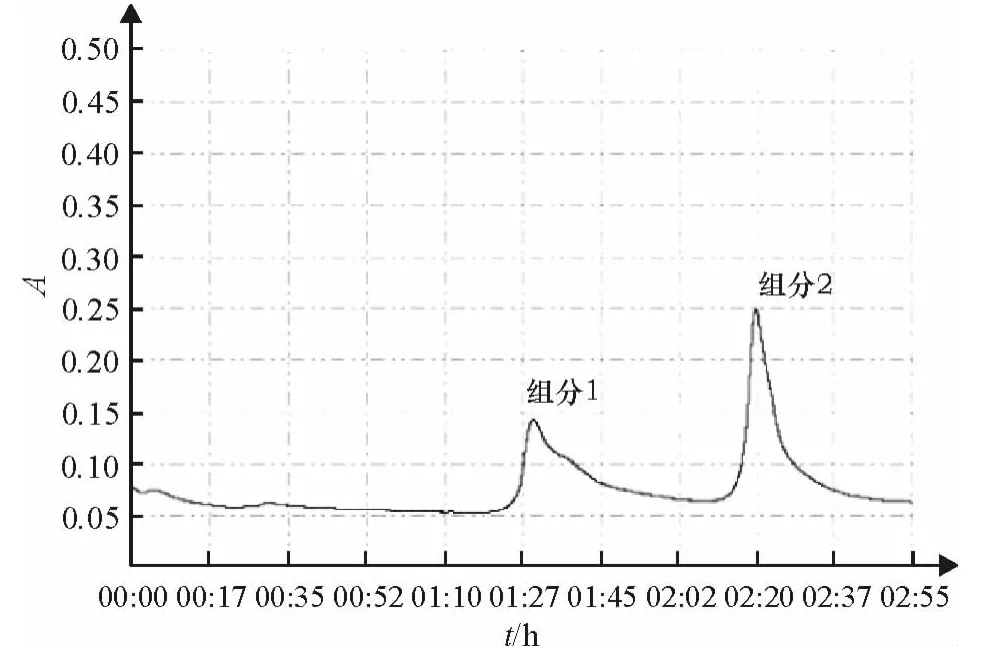
图1 DEAE-纤维素柱的层析图谱Fig.1 The chromatogram of DEAE-cellulose

图2 HAP二次层析的纯化图谱Fig.2 The chromatogram of HAP
(2)经二次层析纯化得到的藻胆蛋白的紫外吸收光谱图见图3.

图3 经二次纯化后的藻胆蛋白的紫外吸收光谱Fig.3 ultraviolet-visible(UV)absorption spectrum of phycobiliprotein
(3)SDS-PAGE电泳结果分析 有文献报道,藻胆蛋白是一类寡聚体蛋白,基本构建单位是α亚基和 β 亚基,分子质量 17 ~22 ku[13],藻红蛋白属于R-型藻红蛋白,除了α和β亚基,还含有γ亚基,α亚基和β亚基的分子质量相当接近,分别为17 ku,18 ku;γ亚基则为37 ku.图4电泳结果显示,藻胆蛋白在17 ku附近有两条带,相当接近,推测这两条带是藻红蛋白的α亚基和β亚基.
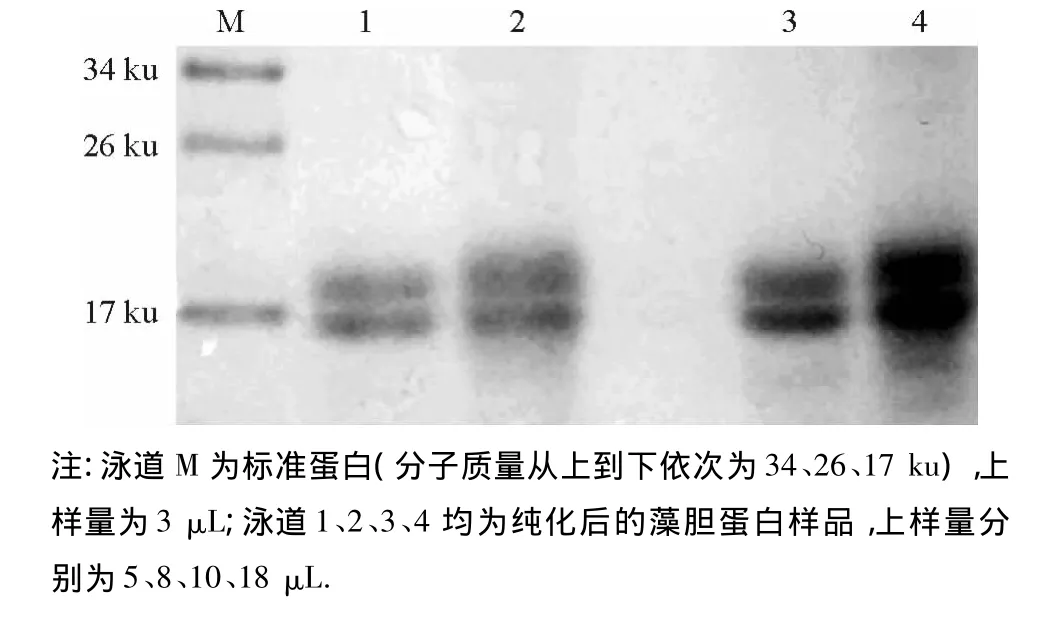
图4 藻胆蛋白的聚丙烯酰胺凝胶电泳图Fig.4 SDS-PAGE picture of phycobiliprotein
2.2 藻胆蛋白对体外培养细胞影响的结果
(1)藻胆蛋白对NIH 3T3细胞生长作用的结果
从实验结果(见图5)可见,与正常细胞对比,藻胆蛋白对NIH 3T3细胞无明显的细胞毒性,并且对NIH 3T3细胞有促增殖的作用,在高质量浓度时(8.65 μg/mL)增殖作用比较明显.

图5 不同质量浓度的藻胆蛋白对NIH 3T3细胞生长作用的影响(n=3)Fig.5 The effect of phycobiliprotein on NIH 3T3 cells(n=3)
(2)NIH 3T3细胞UVA损伤实验造模结果分析倒置显微镜观察显示,与正常细胞对照组比较,NIH 3T3细胞在接受不同剂量UVA辐照后,细胞均受到不同程度的损伤,且NIH 3T3细胞的辐照损伤在强度不变的情况下,与UVA辐射的持续时间存在正相关,随着照射的时间延长,细胞损伤的程度越高.实验结果表明,30 min的UVA辐照时间是细胞半数死亡临界值,是实验造模的理想剂量.
(3)体外抗UVA辐射的实验结果 图6所示,UVA照射,对小鼠NIH 3T3成纤维细胞具有损伤作用,影响细胞的生长;藻胆蛋白具有促进细胞增值的作用,添加藻胆蛋白的细胞生长好于对照组细胞;从UVA损伤+藻胆蛋白组的结果看到,该组的细胞存活率在大于1.08 μg/mL质量浓度后高于对照组,在大于4.32 μg/mL质量浓度后高于藻胆蛋白组,说明藻胆蛋白不但具有促进细胞增殖的作用,而且对UVA损伤的NIH 3T3细胞还具有修复的作用,其中在8.65、4.32、2.16 μg/mL 这3 个质量浓度下的藻胆蛋白对UVA损伤的NIH 3T3细胞修复作用比较明显,细胞的存活率明显高于藻胆蛋白组和对照组.

图6 藻胆蛋白对紫外线损伤NIH 3T3的作用效果Fig.6 The effect of phycobiliprotein on NIH 3T3 exposed to UVA(n=3,*P <0.05;**P <0.01)
(4)细胞周期结果分析 流式细胞技术检测NIH 3T3细胞所处的细胞周期结果显示,藻胆蛋白处理受损的NIH 3T3细胞后,与UVA损伤组相比较,处于G0/G1期的细胞减少,S期的细胞减少,G2/M期的细胞增多,见图7.说明藻胆蛋白使受损伤的NIH 3T3细胞由G1期向S期、G2/M期的转变,细胞DNA合成增长,从而促进NIH 3T3细胞的增殖.
紫外线对生物体的损伤可能通过激活胞膜受体或细胞质的某些活性物质,以及对DNA的直接作用及其miRNA等信号转导通路实现的.本实验的结果显示藻胆蛋白能够保护因UVA辐射损伤的NIH 3T3细胞,激活细胞的修复系统,促进细胞DNA的合成,减少UVA对细胞的伤害.藻胆蛋白是有抗紫外作用的,但是具体的抗紫外机制有待进一步的深入探讨,推测藻胆蛋白的抗紫外作用可能与其抗氧化作用有关联,藻胆蛋白对自由基有很强的清除作用,还能够抑制过氧化物的生成,从而发挥抗氧化作用,减少组织细胞的损伤,从而延缓机体衰老.
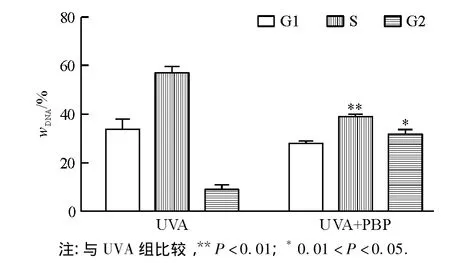
图7 NIH 3T3细胞周期分析结果Fig.7 The cell cycle analysis of NIH 3T3
3 结论
本研究通过超声波破碎提取,饱和硫酸铵盐析分离,DEAE-纤维素和HAP柱层析纯化得到高纯度的藻胆蛋白.通过体外细胞实验,建立了UVA损伤NIH 3T3的细胞模型.紫菜藻胆蛋白不仅可以促进NIH3T3细胞的增殖,而且对UVA损伤的NIH 3T3细胞还有修复的作用.本结果为今后将它作为功能性原料添加到美容护肤产品中的研究开发工作提供基础的理论依据.
藻胆蛋白安全性能好,无毒,能够修复因UVA损伤的成纤维细胞,提高细胞增殖活性,而且与常用的紫外吸收剂相比,藻胆蛋白是天然植物源的活性物质,来源广泛,有很好的可持续的经济意义,因此推断藻胆蛋白在护肤领域应该具有很好的市场价值.
[1]张成武,殷志敏,欧阳平凯.藻胆蛋白的开发与利用[J].中国海洋药物,1995(3):52 -53.
[2]张慧明,王海涛,董银卯,等.紫外线诱导皮肤过敏的损伤类型和机理[J].香料香精化妆品,2010(1):42-45.
[3]王婉如,张 楠,徐俊泉,等.蓝藻中紫外吸收物质的应用研究新进展[J].广州化工,2011,39(17):3-7.
[4]SILVEIRA S T,BURKERT J F M,COSTA J A V,et al.Optimization of phycocyanin extraction from Spirulina platensis using factorial design[J].Bioresource Technology,2007,98(8):1629 -1634.
[5]朱晓君,安辛欣,顾 丽,等.超声辅助同时提取条斑紫菜多糖及藻胆蛋白工艺的优化[J].食品科学,2008,29(5):241 -244.
[6]肖海芳,马海乐,朱文学,等.紫菜中蛋白和多糖的脉冲超声辅助提取工艺研究[J].食品与机械,2006,22(5):60-63.
[7]蔡春尔,周 铭,李春霞,等.条斑紫菜藻胆蛋白提纯方法优化探索[J].生物技术通报,2009(2):98-102.
[8]程 超,薛 峰,汪兴平,等.藻胆蛋白提取纯化及生理活性研究进展[J].食品科学,2012,33(9):251-259.
[9]MORAES C C,KALIL S J.Strategy for a protein purification design using C-phycocyanin extract[J].Bioresource Technology,2009,100(21):5312 -5317.
[10]李春霞,吴淑贤,蔡春尔,等.条斑紫菜藻红、藻蓝蛋白逐级放大的纯化工艺[J].中国生物工程杂志,2010,30(1):67-72.
[11]罗振宇.干细胞分泌上清的制备及抗氧化和抗皮肤光老化活性研究[D].广州:暨南大学,2010.
[12]HELGA MERWALD,GABRIELE KLOSNER,CLAUDIA KOKESCH,et al.UVA-induced oxidative damage and cytotoxicity depend on the mode of exposure[J].Photochemistry and Photobiology B:Biology,2005,79:197 -207.
[13]王庭健,林 凡,赵方庆,等.藻胆蛋白及其在医学中的应用[J].植物生理学通讯,2006,42(2):303-307.
猜你喜欢
昆明医科大学学报(2021年8期)2021-08-13
云南医药(2021年3期)2021-07-21
孩子(2021年1期)2021-02-02
世界最新医学信息文摘(2020年68期)2020-12-25
烟台大学学报(自然科学与工程版)(2020年1期)2020-02-08
幽默大师(2019年10期)2019-10-17
婚姻与家庭·婚姻情感版(2018年4期)2018-04-09
中国现代医学杂志(2015年26期)2015-12-23
小朋友·快乐手工(2015年6期)2015-07-01
中国当代医药(2015年33期)2015-03-01
