
不同海拔下烤烟叶片全展后光合特性研究
2013-10-16 11:15江厚龙张正扬
植物营养与肥料学报 2013年6期
江厚龙, 王 瑞, 贾 峰, 张 均, 张正扬
(1 重庆烟草科学研究所,重庆 400715; 2 湖北省烟草公司恩施州公司,湖北恩施 445000; 3 河南工业大学生物工程学院,河南郑州 450001; 4 河南科技大学,河南洛阳 471003; 5 江西省烟草公司,江西南昌 330025)
不同海拔下烤烟叶片全展后光合特性研究
江厚龙1, 王 瑞2*, 贾 峰3, 张 均4, 张正扬5
(1 重庆烟草科学研究所,重庆 400715; 2 湖北省烟草公司恩施州公司,湖北恩施 445000; 3 河南工业大学生物工程学院,河南郑州 450001; 4 河南科技大学,河南洛阳 471003; 5 江西省烟草公司,江西南昌 330025)
在湖北省恩施州选择同一山脉的3个不同海拔点(500 m、 900 m、 1300 m)进行盆栽试验,研究了3个不同基因型烤烟品种K326、 云烟87、 中烟103在不同海拔下烤烟叶片全展后的光合特性。结果表明,同一海拔下,各品种烤烟的净光合速率(Pn)在叶片全展时达到最高,随后逐渐下降,随着海拔升高,烤烟叶片光合功能衰退减慢,光合功能期延长,光合生产能力增加。RuBPCase初始活性下降是导致烤烟叶片全展后Pn下降的主要因素。烤烟叶片全展后叶绿体结构逐步解体,且海拔越高开始解体的时间越晚。供试三个品种,以中烟103在试验各海拔点均表现出较高的光合生产能力,为高光效品种。
海拔; 烤烟; 光合特性
Abstract: Pot experiments were conducted at the altitudes of 500 m, 900 m and 1300 m in Enshi Autonomous Prefecture of Hubei to study the differences of photosynthetic characteristics of flue-cured tobacco cultivars with genotypes K326, Yunyan87 and Zhongyan103. The result show that the net photosynthetic rates(Pn) are maximum at full expansion of leaves of the flue-cured tobacco at various altitudes in Enshi, then decrease gradually, which is non-stomatal limitation that causes the decrease ofPn. With the increase of planting altitude, photosynthetic function of leaves is declined more slowly, the photosynthetic function period is prolonged, and the photosynthetic capacity of leaves is raised. Compared with the change of the chlorophyll content, the decrease of activities of RuBPCase is the main factor of the decline of photosynthetic function in tobacco leaves. The chloroplast microstructure is disintegrated gradually after the full expansion of leaves, and with the rising of the altitude, the start time of descending of chloroplast microstructure is delayed. The photosynthetic efficiency of the Zhongyan103 cultivar is highest because of the higher photosynthetic capacity in various elevations.
Keywords: elevation; flue-cured tobacco; photosynthetic characteristic
光合作用是作物产量和质量形成的基础,作物中90%以上的干物质直接或间接来自光合作用。烟草产量和品质的形成依赖于光合作用产生的有机物质,提高烟草产量和品质的根本途径是改善烟草的光合性能[1]。海拔高度涉及到了温度、 湿度、 光照等诸多环境因子,是影响植物生长发育、 物质代谢、 结构和功能等重要的生态因素之一[2]。海拔综合了大多数影响作物光合作用的因子,不同的海拔代表着不同的生态环境系统,因此,海拔高度给我们研究整体生态系统对作物光合作用的影响提供了一个理想的区域。
关于海拔对植物光合特性影响的研究较多,但由于不同植物自身生理特性的差异及研究区域不同,其研究结果也不尽一致。祖元刚等[3]以青藏高原的裂叶沙参为研究对象,其结果表明,低海拔的裂叶沙参的净光合速率(Pn)和表观量子效率(AQY)比高海拔地区低;刘华等[4]利用天山雪莲研究表明,净光合速率和最大净光合速率随海拔的升高而降低,其光能利用率却与此相反。Kumar等[5-6]在对大麦、 小麦以及豌豆的研究表明,不同海拔间的Pn没有明显差异,高海拔地区有较高的气孔导度、 蒸腾速率以及较低的水分利用率;高海拔和低海拔RuBP羧化酶的初始活力没有显著差异,但高海拔RuBP羧化酶的活性比低海拔的低。
本文以3个遗传背景不同的烤烟品种为材料,采用盆栽试验方法,对不同海拔下烤烟叶片全展后光合特性进行研究,旨在探讨不同海拔生态系统对烤烟光合性能影响及其形成的生理生化机理,明确不同品种烤烟生态适应性,为进一步丰富烟草栽培生理理论及进行烟草种植区划提供科学参考。
1 材料与方法
1.1 试验设计
试验于2009年在湖北省恩施州宣恩县同一山脉的3个不同海拔点进行,分别为海拔500 m(109°22′8″E、 29°57′5″N)、 海拔900 m(109°22′45″E、 29°58′15″N)和海拔1300 m(109°23′13″E、 29°59′5″N)。采用盆栽试验,各海拔点分别种植K326、 云烟87和中烟103三个烤烟品种,每品种栽烟20盆(盆高40 cm,直径35 cm,装干土15 kg),共60盆,按行、 株距110 cm×50 cm排成3排,各品种完全随机排列。采用同一土壤,土壤类型为黄棕壤,质地偏粘,pH值为6.73,含有机质36.6 g/kg、 碱解氮184.62 mg/kg、 速效磷35.53 mg/kg、 速效钾221.0 mg/kg。每株施纯氮量5 g,氮、 磷、 钾比例为1 ∶1.5 ∶2.5,所有肥料均作基肥一次性施入。各试验点搭建简易遮雨棚,下雨时进行遮盖,完全人工灌溉,保证各试验点土壤水分的一致性。各点均采用漂浮育苗,统一于5月12日移栽,其它管理均按常规措施进行。
各品种选择代表性烟株5株,每株标记中部叶(自下而上第9位叶),观察叶片全展时期。同一海拔各品种达到全展时间基本一致,而海拔500 m在移栽后45 d达到全展,海拔900 m在移栽后50 d达到全展,海拔1300 m在移栽后58 d达到全展。
1.2 测定项目和方法

1.2.2 叶绿素含量 如1.2.1所述时间,采用丙酮、 乙醇混合液法[7]测定叶绿素含量。
1.2.3 RuBP羧化酶初始活性 在叶片全展时、 全展后14 d和全展后28 d,取鲜样迅速用液氮进行保存。叶片加酶提取液(50 mmol/L Tris-HCl,pH8.0,1 mmol/L EDTA-Na2,5 mmol/L DTT,1 mmol/L PMSF,PVP少许)冰浴研磨,12000×g离心15 min,上清液即为RuBPCase粗提液。在含有NaHCO3的0.5 mL反应介质中加入10 μL酶粗提液,反应45 s后加入2 μmol/L的HCl终止反应。反应后的混合液于80℃烘干,加入0.5 mL蒸馏水使烘干物充分溶解,再加入4.5 mL闪烁液,用液体闪烁计数仪测定放射强度,换算出酶活性[8]。
1.2.4 叶绿体超微结构 如1.2.1所述时间,以烤烟云烟87为材料,观测不同海拔下烤烟叶片细胞超微结构的变化。具体方法为: 用去离子水洗净,棉布拭干,在中脉附近取叶肉组织,用刀片切成1 mm×2 mm小片,后将切好的组织块迅速投入2.5%戊二醛固定液中,抽气至完全下沉,4℃下保存,后用磷酸缓冲液(pH 7.6)洗涤3次,每次20 min。再将该材料转移到1%锇酸中,置4℃下后固定4 h,蒸馏水洗涤3次,每次20 min;随后经系列乙醇脱水,环氧树脂618包埋,LKB-V型超薄切片机(瑞典LKB公司)切片,醋酸双氧铀和柠檬酸铅双重染色后于Hitach iH-600透射电镜下观察并拍照[9]。
1.2.5 相关光合功能指标的计算
净光合速率高值持续期(Active photosynthetic duration,APD): 叶片全展至叶片下降至全展时的50%所持续的时间[10]。
叶绿素含量缓降期(Relative steady phase of chlorophyll content, RSP): 叶片全展至叶绿素含量下降至全展时的80%所持续的时间[11]。
光合叶面积(Leaf area,LA): 用数码相机和Photoshop软件计算每张叶片的叶面积[12]。
叶源量(Leaf source capacity,LSC): 指单个叶片在整个生育期内同化CO2的总量,按以下公式[13]计算:
式中,Pn为净光合速率,D为测定间隔时间,LA为单叶全展时的面积,m为测定次数,i=1, 2, …, m
1.3 统计分析
所有数据均用Excel录入数据、 制作图表和计算,用DPS[14]软件进行相关统计分析。
2 结果与分析
2.1 净光合速率(Pn)
从图1可见,不同海拔下各品种叶片Pn均在全展时最高,随着测定时间推迟而下降,测定时期间差异均达到极显著水平(P<0.01)。表明叶片全展后,烤烟叶片光合功能不断衰退。K326在海拔500、 900和1300 m的Pn下降率分别为84.8%、 82.9%和75.6%,云烟87分别为85.4%、 81.2%和70.48%,中烟103分别为70.6%、 63.8%和59.6%,均表现为随着海拔升高而减少的趋势。这表明海拔越高烤烟叶片光合能力衰退越慢。比较各品种在全展时、 全展后14 d和全展后28 d海拔间变异系数(CV)和最大差异率(PR)可以看出,随着测定时间推迟CV和PR均呈增大趋势(表1)。这表明随着叶片生育进程的推进,海拔对烤烟叶片Pn影响逐渐增大。

图1 不同海拔下烤烟叶片全展后净光合速率的变化Fig.1 Changes of Pn after full expansion of leaves of flue-cured tobacco at different altitudes

品种Cultivar项目Item净光合速率Pn0d14d28d叶绿素Chla+b0d14d28dRuBP羧化酶初始活性RuBPCaseinitalactivity0d14d28dK326变异系数CV(%)1.98.225.88.110.714.03.87.928.6最大差异率PR(%)4.322.180.420.228.641.29.821.4109.1云烟87Yunyan87变异系数CV(%)0.68.627.52.58.817.91.31.630.2最大差异率PR(%)1.523.683.56.323.454.22.94.166.7中烟103Zhongyan103变异系数CV(%)0.96.018.22.93.06.63.04.511.3最大差异率PR(%)2.315.852.67.47.517.67.411.530.4
2.2气孔导度(Gs)和胞间CO2浓度(Ci)
从图2可见,不同海拔下各品种烤烟叶片Gs均随着测定时间的推迟表现先增加后降低的趋势。在海拔500 m和900 m,Gs均在全展后7 d达到峰值;在海拔1300 m,Gs均在叶片全展后14 d达到峰值。不同海拔下各品种烤烟叶片胞间CO2浓度(Ci)均随着测定时间的推迟而呈上升的趋势(图3),和净光合速率Pn的变化相反。在叶片全展后7 d或14 d内,Pn下降伴随着Gs增加;在叶片全展后7天或14天后,Pn下降,Gs也下降但伴随Ci增加。根据Farqhar和Sharkey的观点,可以推测烤烟叶片全展后Pn下降的主要原因是非气孔限制,即叶肉细胞光合活性的下降引起[15]。

图2 不同海拔下烤烟叶片全展后气孔导度的变化Fig.2 Changes of Gs after full expansion of leaves of flue-cured tobacco at different altitudes
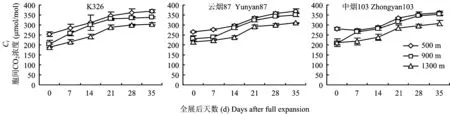
图3 不同海拔下烤烟叶片全展后胞间CO2浓度的变化Fig.3 Changes of Ci after full expansion of leaves of flue-cured tobacco at different altitudes
2.3 叶绿素含量
从图4可见,各品种不同海拔处理叶片叶绿素(Chl a+b)含量均随着测定时间的推移而下降,测定时期间差异均达到极显著水平(P<0.01)。各品种在海拔500、 900和1300 m的Chl a+b含量平均值均表现随着海拔升高而增加趋势,海拔间差异均达到显著水平(P<0.05)。K326在海拔500、 900和1300 m Chl a+b含量的下降率分别为67.6%、 60.1%和55.2%;云烟87的下降率分别为65.1%、 53.3%和46.5%;中烟103的下降率分别为64.0%、 60.0%和59.2%,均表现为随着海拔的升高下降率呈减少趋势,表明海拔越高烤烟叶片叶绿素降解越慢。从表1可以看出,各品种Chl a+b含量海拔间变异系数和最大差异率均随着测定时间的推迟呈增大趋势,表明随着叶片生育进程的推进,海拔对烤烟叶片叶绿素含量影响逐渐增大。
2.4 RuBP羧化酶初始活性
从图5可见,不同海拔下各品种烤烟叶片RuBP羧化酶(RuBPCase)初始活性均随着测定时间的推迟而下降,各时期间差异达到极显著水平(P<0.01)。在叶片全展时,各品种不同海拔间差异均未达到显著水平(P>0.05);在叶片全展后14 d,K326表现出随海拔升高而增加的趋势,海拔间差异达到显著水平(P<0.05),而云烟87和中烟103不同海拔间差异未达到显著水平(P>0.05);在叶片全展后28 d,各品种在海拔900 和1300 m之间差异未达到显著水平(P>0.05),它们均显著高于海拔500 m(P<0.05)。K326在海拔500、 900和1300 m的RuBPCase初始活性下降率分别为82.81%、 68.65%和62.29%,云烟87分别为78.87%、 69.01%和63.76%,中烟103分别为68.05%、 57.35%和58.9%,均表现出随海拔的升高而下降的趋势,表明海拔越高光合酶活性衰退越慢。从表1可以看出,各品种RuBPCase初始活性海拔间变异系数和最大差异率均随着测定时间的推迟呈增大趋势,表明随着叶片生育进程的推进,海拔对烤烟叶片RuBPCase初始活性的影响逐渐增大。

图4 不同海拔下烤烟叶片全展后叶绿素含量的变化 Fig.4 Changes of content of Chl a+b after full expansion of leaves of flue-cured tobacco at different altitudes
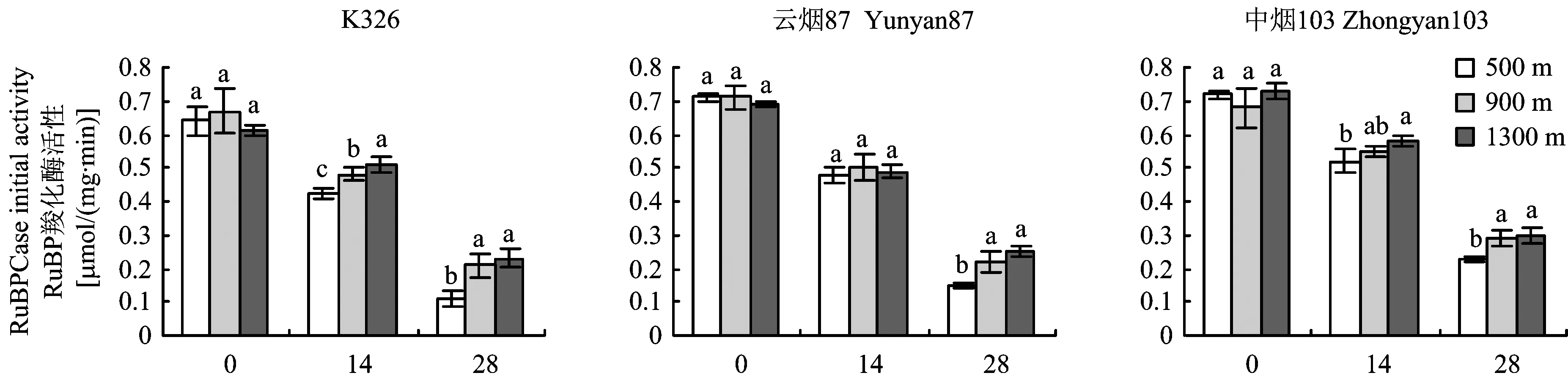
图5 不同海拔下烤烟叶片全展后RuBP羧化酶的初始活性的变化Fig.5 Changes of RuBPCase initial activity after full expansion of leaves of flue-cured tobacco at different altitudes
2.5 叶绿体超微结构
2.6 相关光合功能指标比较
统计分析结果显示(表2),净光合速率(Pn)均值和净光合速率高值持续期(APD)在海拔间和品种间差异均达到极显著水平(P<0.01);叶绿素含量缓降期(RSP)在海拔间和品种间差异均不显著(P>0.05);叶源量(LSC)在海拔间达到极显著水平(P<0.01),但在品种间差异不显著(P>0.05);海拔和品种互作对Pn均值影响未达到显著水平,但对其他各指标的影响均达到极显著水平(P<0.01)。
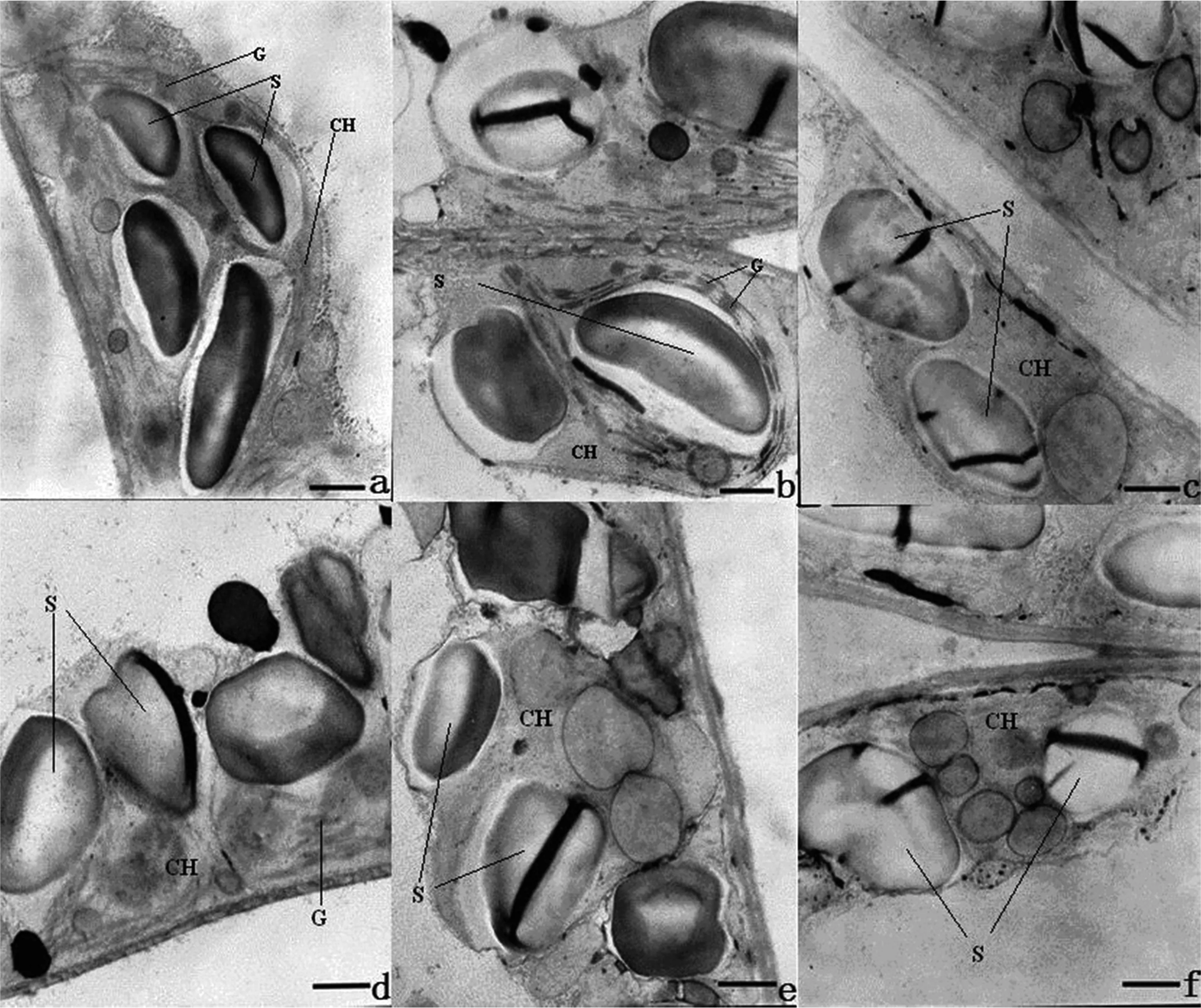
图6 海拔500 m处烤烟叶片全展后叶绿体超微结构的变化Fig.6 Variation in ultrastructure of chloroplast after full expansion of leaves of flue-cured tobacco at 500 m[注(Note): a、 b、 c、 d、 e、 f 依次为叶片全展后0 d、 7 d、 14 d、 21 d、 28 d、 35 d. a, b, c, d, e and f are 0 d, 7 d, 14 d, 21 d, 28 d and 35 d after full expansion of leaves.×10K,Bar=1μm. CH—叶绿体Chloroplast, S—淀粉粒Starch grain, G—基粒类囊体Granum thylakoid.]
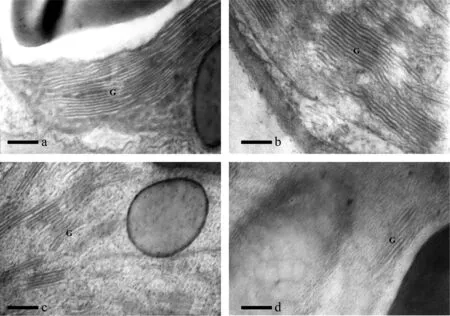
图7 海拔500 m处烤烟叶片全展后类囊体片层结构的变化Fig.7 Variation in ultrastructure of thylakoid lamellar after full expansion of leaves of flue-cured tobacco at 500 m[注(Note): a、 b、 c、 d 依次为叶片全展后0 d、 7 d、 14 d、 21 d. a, b, c and d are 0 d, 7 d, 14 d and 21 d after full expansion of leaves. ×40K,Bar=250 nm. G—基粒类囊体Granum thylakoid.]

图8 海拔900 m处烤烟叶片全展后叶绿体超微结构的变化Fig.8 Variation in ultrastructure of chloroplast after full expansion of leaves of flue-cured tobacco at 900 m[注(Note): a、 b、 c、 d、 e、 f 依次为叶片全展后0 d、 7 d、 14 d、 21 d、 28 d、 35 d. a, b, c, d, e and f are 0 d, 7 d, 14 d, 21 d, 28 d and 35 dafter full expansion of leaves. ×10K,Bar=1μm,CH—叶绿体Chloroplast, S—淀粉粒Starch grain, G—基粒类囊体Granum thylakoid.]
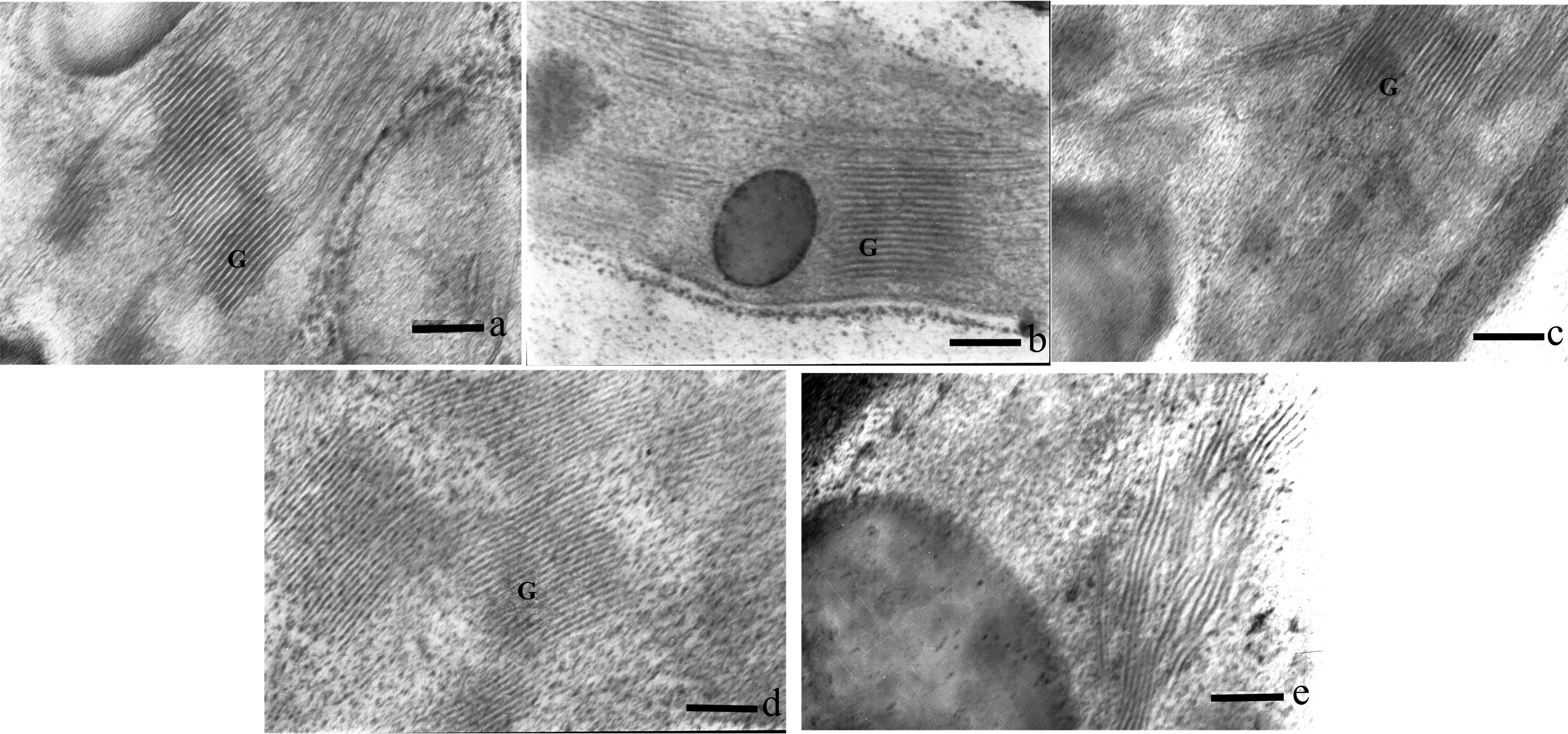
图9 海拔900 m处烤烟叶片全展后类囊体片层结构的变化Fig.9 Variation in ultrastructure of thylakoid lamellar after full expansion of leaf in flue-cured tobacco at 900 m[注(Note): a、 b、 c、 d、 e依次为叶片全展后0 d、 7 d、 14 d、 21 d、 28 d. a, b, c, d and e are 0 d, 7 d, 14 d, 21 d and 28 d after fullexpansion of leaves. ×40K, Bar=250 nm. G—基粒类囊体Granum thylakoid.]
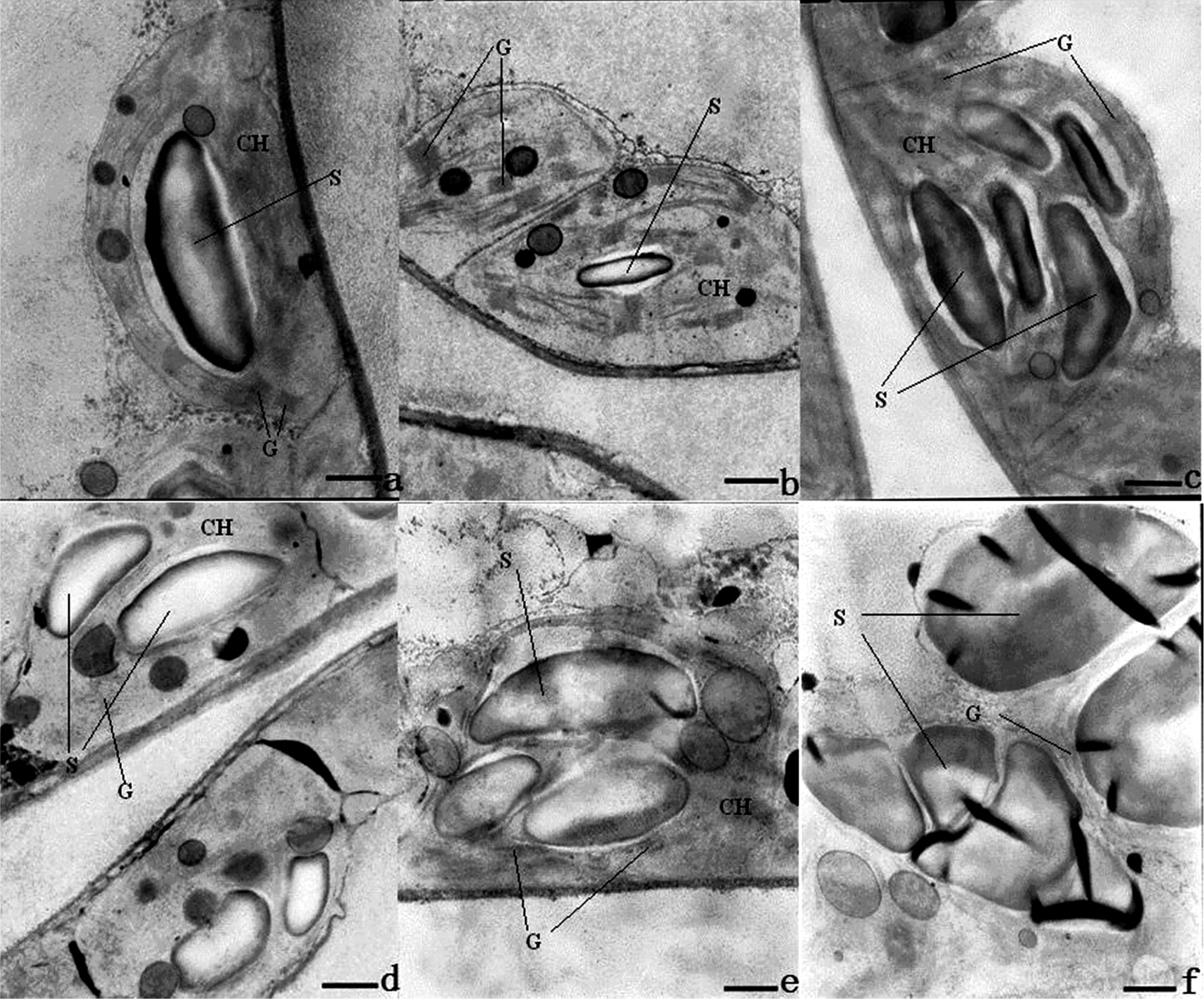
图10 海拔1300 m处烤烟叶片全展后叶绿体超微结构的变化Fig.10 Variation in ultrastructure of chloroplast after full expansion of leaves of flue-cured tobacco at 1300 m[注(Note): a、 b、 c、 d、 e、 f 依次为叶片全展后0 d、 7 d、 14 d、 21 d、 28 d、 35 d. a, b, c, d, e and f are 0 d, 7 d, 14 d, 21 d, 28 d and 35 d after full expansion of leaves. ×10K, Bar=1μm,CH—叶绿体Chloroplast, S—淀粉粒Starch grain, G—基粒类囊体Granum thylakoid.]
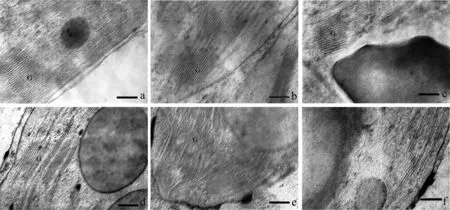
图11 海拔1300 m处烤烟叶片全展后类囊体片层结构的变化Fig.11 Variation in ultrastructure of thylakoid lamellar after full expansion of leaf in flue-cured tobacco at 1300 m[注(Note): a、 b、 c、 d、 e、 f 依次为叶片全展后0 d、 7 d、 14 d、 21 d、 28 d、 35 d. a, b, c, d, e and f are 0 d, 7 d, 14 d, 21 d, 28 d and 35 dafter full expansion of leaves.×40K, Bar=250 nm. G—基粒类囊体Granum thylakoid.]

项目ItemFPnFAPDFRSPFLSC品种间Cultivar25.81**27.50**1.59665.90海拔间Altitude86.06**103.54**3.779724.51**品种×海拔Cultivar×Altitude0.68450.64**4173.477**1530.90**
注(Note): APD—Active photosynthesis duration, RSP—Relative steady phase of chlorophyll content, LSC—Leaf source capacity.FPn、FAPD、FRSP、FLSC依次表示Pn平均值、 APD、 RSP和LSC的F检验值. **表示达到0.01极显著水平.FPn,FAPD,FRSPandFLSCareFtest values of averagesPn, APD, RSP and LSC, respectively. ** indicates the 0.01 significant level.
进一步分析表明(表3),各品种叶片Pn平均值、 APD和LSC均表现出随着海拔升高而增加趋势,不同海拔间差异达到显著水平(P<0.05)。表明随着海拔升高烤烟叶片的光合功能期延长,光合生产能力增加。在各个海拔下,均以中烟103 的Pn均值、 APD和LSC最高,显著高于其他两个品种,说明中烟103具有较长的光合功能期和较高光合生产能力。在海拔900 m,K326和云烟87的Pn平均值、 APD和LSC差异不显著。在海拔500 m和1300 m,K326和云烟87的Pn平均值差异不显著,但K326的APD和LSC均显著低于云烟87。表明K326在海拔900 m外的其他区域光合功能期较短,光合生产能力较低。与APD相比,RSP各处理间差异没有表现出明显规律。
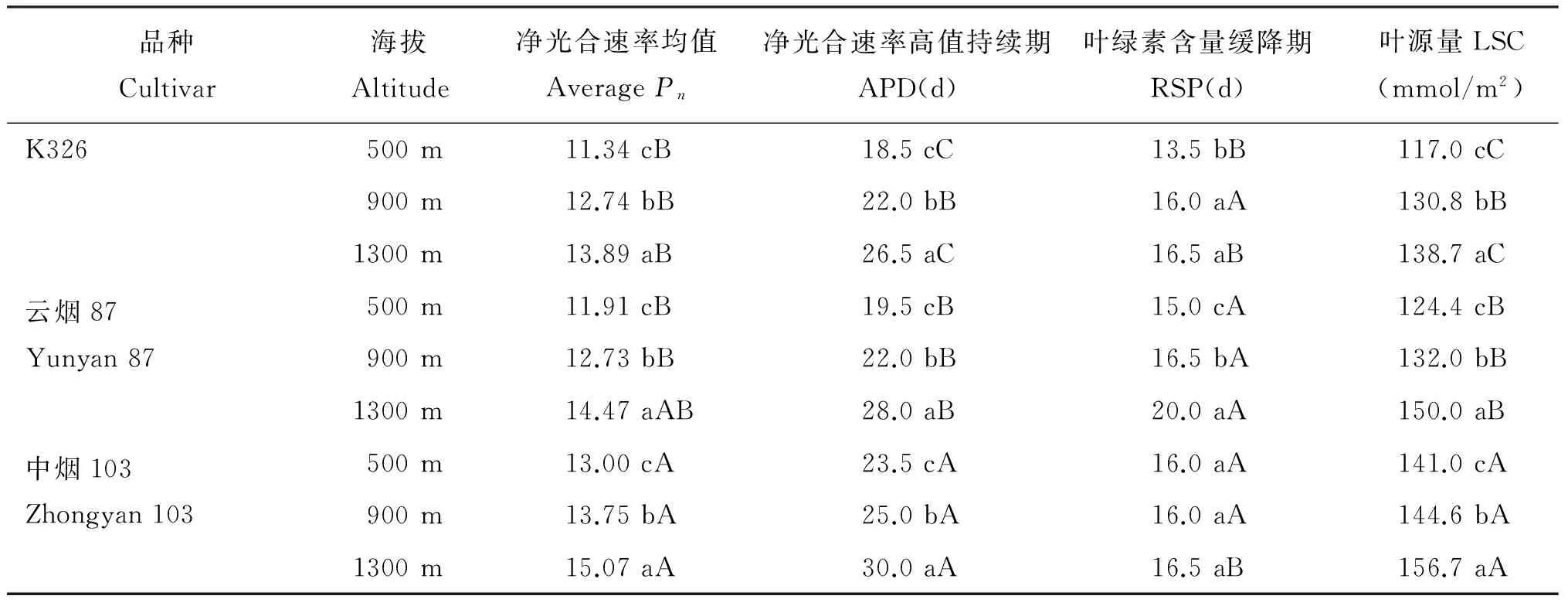
表3 不同海拔下烤烟叶片的Pn均值、 APD、 RSP和LSC
注(Note): APD—Active photosynthetic duration, RSP—Relative steady phase of chlorophyll content, LSC—Leaf source capacity.同列数据后不同小写字母表示同一品种不同海拔高度间在0.05水平差异显著 Values followed by different small letters within a column are significantly different among different altitudes within a cultivar at 0.05 level. 同列数据后不同大写字母表示同一海拔高度下不同品种间在0.05水平差异显著 Values followed by different capital letters within a column are significantly different amont different cultivars within a altitude at 0.05 level.
3 讨论
从环境角度分析可知,处于不同环境条件下植株光合功能衰退的程度有较大差异。本研究表明,海拔越高烤烟叶片光合功能衰退越慢。恩施州烤烟的成熟阶段正处于炎热的夏季,海拔500 m、 900 m和1300 m处测定时的环境温度均值分别为29.1、 26.1和24.7℃,叶片全展后7天日均温分别为35.6℃、 34.1℃和29.7℃,海拔越低环境温度越高。高温会影响植物的代谢和调节过程,导致植物生理功能衰退[22],从而加速了光合功能的衰退。同时因为低海拔区域前期较高的土壤温度加快了土壤养分转化的速度,加快了烟株的生长,消耗了大量的养分而导致后期养分的缺乏,从而表现出随着叶片生育进程推进,海拔对烤烟叶片光合能力影响逐渐增大。
叶片的衰老导致细胞结构和新陈代谢变化,其中叶绿体是叶片衰老的一个重要器官。叶绿体的变化分为两个阶段,首先是叶绿体的形态异常;然后,随着非正常化加剧而逐渐消失[23]。从本研究来看,海拔500、 900和1300 m处烤烟叶片细胞叶绿体结构发生明显变化的时期分别在叶片全展后14 d、 21 d和28 d,表现出随着海拔的升高而推迟的趋势,这与不同海拔下光合功能衰退程度的变化趋势是一致的。通常认为,叶绿体类囊体的垛叠程度是决定其光合能力的主要因子[24];张洁等[25]在对番茄的研究中也指出,净光合速率与基粒片层的垛叠程度密切相关。烤烟叶片在全展时叶绿体基粒类囊片层最为发达,因此最有利于光能的吸收和转化。不同海拔下烤烟叶片类囊片层减少的程度不一致,从而引起光合功能下降的程度不一致,低海拔地区在叶片全展后21 d类囊片层就基本消失,而高海拔地区在全展后35 d还可以观察到少量的类囊片层。这就从解剖生理学角度解释了不同海拔下烤烟叶片光合能力的差异。
关于海拔对作物光合能力影响的研究前人开展了很多,本研究将前人[26]在评价小麦、 水稻等作物光合能力时所用指标如叶源量(LSC)、 光合速率高值持续期(APD)、 叶绿素含量相对稳定期(RSP)等应用到烟草中,更加全面系统地评价不同海拔下3个品种烤烟光合生产能力的差异。总体看来,生长在不同海拔下的烤烟叶片光合生产能力存在着显著差异,随着海拔的升高,光合功能期逐渐延长,光合生产能力逐渐增加。不同品种烤烟的光合生产能力也存在着显著差异,在参试的3个品种中,中烟103在各个海拔点均表现出较高的光合生产能力,从这个角度来说,中烟103是一个高光效品种。本研究还发现,同一处理RSPAPD并不相近,RSP明显要低于APD,这与江力等[27]关于NC89和JYH两个品种烟草叶片RSP和APD极为接近的结论不一致,其原因有待进一步研究。
[1] 韩锦峰. 烟草栽培生理学[M]. 北京: 中国农业出版社, 2003 Han J F. Tobacco Cultivation[M]. Beijing: China Agriculture Press, 2003.
[2] Woodward F I. Ecophysiological studies on the shrubVacciniummyrtillusL. taken from a wide altitudinal range[J]. Oecologia, 1986, 70: 580-586.
[3] 祖元刚, 阎秀峰, 张文辉, 等. 青藏高原东部不同海拔高度裂叶沙参气体交换与水分利用效率[J]. 植物学报, 1998, 40(10): 947-954. Zu Y G, Yan X F, Zhang W H,etal. Gas exchange and water use efficiency ofAdenophoraLobophYlla(Campanulaceae) at different altitudes on the east boundary of the Qing-Zang Plateau[J]. Acta Bot. Sin., 1998, 40(10): 947-954.
[4] 刘华, 臧润国, 张新平, 等. 天山中部3种自然生境下天山雪莲的光合生理生态特性[J]. 林业科学, 2009, 45(3): 40-47. Liu H, Zang R G, Zhang X Petal. Photosynthetic and ecophysiological characteristics of snow lotus(Saussueainvolucrata) in its natural conditions in the central part of Tianshan Mountains[J]. Sci. Silv. Sin., 2009, 45(3): 40-48.
[5] Kumar N, Kumar S, Ahuja P S. Photosynthetic characteristics ofHordeum,Triticum,Rumex, andTrifoliumspeciesat contrasting altitudes[J]. Photosynthetica, 2005, 43(2): 195-201.
[6] Kumar N, Kumar S, Ahuja P S. Differences in the activation state of ribulose-1, 5-bisphosphate carboxylase/oxygenase in barley, pea, and wheat at two altitudes[J]. Photosynthetica, 2004, 42(2): 303-305.
[7] 张宪政. 植物叶绿素含量测定—丙酮乙醇混合液法[J]. 辽宁农业科学, 1986, (3): 26-28. Zhang X Z. Plant chlorophyll determination acetone ethanol mixture method[J]. Liaoning Agric. Sci., 1986, (3): 26-28.
[8] 宁正祥, 李明启. 变酸显色法同时测定核酮糖1, 5二磷酸羧化酶加氧酶活性[J]. 植物生理学报, 1990, 16(2): 131-138. Ning Z X, Li M Q. A colorimetric method for simultaneous determination of ribulose-1, 5-bisphosphate carboxylase/oxygenase activities using chromotropic acid[J]. Acta Phytophysiol. Sin., 1990, 16(2): 131-138.
[9] 张华, 翁梦苓, 梁志敏. 烟草腺毛发育过程中叶绿体形态学研究[J]. 西北植物学报, 2008, 28(8): 1592-1595. Zhang H, Wen M L, Liang Z M. Morphological research on chloroplast of tobacco trachoma during development[J]. Acta Bot. Bor. Occid. Sin, 2008, 28(8): 1592-1595.
[10] 张荣铣, 程在全, 方志伟等. 关于光合速率高值持续期的初步研究[J]. 南京师范大学学报, 1992, 15: 76-86. Zhang R X, Cheng Z Q. Preliminary study on the net photosynthetic active duration of leaf in wheat[J]. J. Nanjing Normal Univ.(Nat. Sci. Ed.), 1992, 15: 149-1601.
[11] 曹树青, 翟虎渠, 张红生等. 不同类型水稻品种叶源量及有关光合生理指标的研究[J]. 中国水稻科学, 1999, 13(2): 91-94. Cao S Q, Cui H Q, Zhang H S Hetal. Leaf source capacity and photosynthetic indexes in different types of rice varieties[J]. Chin. J Rice Sci., 1999, 13(2): 91-94.
[12] 肖强, 叶文景, 朱珠, 等. 利用数码相机和photoshop 软件非破坏性测定叶面积的简便方法[J]. 生态学杂志, 2005, 24(6): 711-714. Xiao Q, Ye W J, Zhu Zetal. A simple non-destructive method to measure leaf area using digital camera and Photoshop software[J]. Chin. J. Ecol., 2005, 24(6): 711-714.
[13] 张永丽, 肖凯, 李雁鸣. 种植密度对杂种小麦C6-38/Py85-1旗叶光合特性和产量的调控效应及其生理机制[J]. 作物学报, 2005, 31(4): 498-505. Zhang Y L, Xiao K, Li Y M. Effects and physiological mechanism of planting densities on photosynthesis characteristics of flag leaf and grain yield in wheatHybridC6238PPy8521[J]. Acta Agon. Sin., 2005, 31(4): 498-505.
[14] Tang Q Y, Feng M G. DPS data processing system: experimental design, statistical analysis, and data mining[M]. Beijing: Science Press, 2007. 82-173.
[15] Farqhar G D, Sharkey T D. Stomatal conductance and photosynthesis[J]. Ann. Rev. Plant Physiol., 1982, 33: 317-345.
[16] 匡廷云, 卢从明, 李良璧,等. 作物光能利用效率与调控[M]. 济南: 山东科学技术出版社,2004: 117-118. Kuang T Y, Lu C M, Li L Betal. Photosynthetic efficiency of crops and regulation[M]. Jinan: Shandong Science and Technology Press, 2001: 117-118.
[17] 刘国顺. 烟草栽培学[M]. 北京: 中国农业出版社, 2003. Liu G S. Tobacco cultivation[M]. Beijing: China Agricultural Press, 2003
[18] 许大全.光合作用效率[M]. 上海: 上海科学技术出版社, 2002: 86-95. Xu D Q. Photosynthesis efficiency[M]. Shanghai: Shanghai Science and Technology Press, 2002: 86-95.
[19] 许大全. 光合作用气孔限制分析中的一些问题[J]. 植物生理学通讯, 1997, 33(4): 241-244. Xu D Q. Some problems in stomatal limitation analysis of photosynthesis[J]. Plant Physiol. Comm., 1997, 33(4): 241-244.
[20] 王婵娟, 刘国顺, 张彩霞. 烟叶生长发育过程中叶片光合特性及叶绿体超微结构的变化[J]. 江西农业大学学报, 2010, 32(2): 254-259. Wang C J, Liu G S, Zhang C X. Changes in photosynthetic characteristics and chloroplast ultrastructure during the development of tobacco leaf[J]. Acta Agric. Univ. Jiangxiensis, 2010, 32(2): 254-259.
[21] 蒋德安, 陆庆, 翁晓燕, 等. 水稻剑叶衰老期Rubisco活化酶对Rubisco活力和光合速率的调节[J]. 浙江大学学报(农业与生命科学版), 2000, 26(2): 119-124. Jiang D A, Lu Q, Wen X Y. Regulation of Rubisco carboxylation activity and photosynthetic rate by Rubisco activase during leaf senescence in rice[J]. J. Zhejiang Univ.(Agric. Life Sci.), 2000, 26(2): 119-124.
[22] 李俊清, 牛树奎. 森林生态学[M]. 北京: 高等教育出版社, 2006. Li J Q. Niu S K. Forest ecology[J]. Beijing: Higher Education Press, 2006.
[23] Ford D M, Shibles R. Photosynthesis and other traits in relation to chloroplast number during soybean leaf senescence[J]. Plant Physiol., 1988, 86: 108-111.
[24] 李寒冰, 胡玉熹, 白克智, 等. 小麦芒和旗叶叶绿体结构及低温荧光发射光谱的比较研究[J]. 电子显微学报, 2002, 21(2): 97-101. Li H B, Hu Y X, Bai K Zetal. Comparison of chloroplast ultrastructure and 77K fluorescence emission spectra between awns and flag leaves in wheat[J]. J. Chin. Electr. Microsc. Soc., 2002, 21(2): 97-101.
[25] 张洁, 李天来. 日光温室亚高温对番茄光合作用及叶绿体超微结构的影响[J]. 园艺学报, 2005, 32(4): 614-619. Zhang J, Li T L. Effects of daytime sub-high temperature on photosynthesis and chloroplast ultra structure of tomato leaves in greenhouse[J]. Acta Hort. Sin., 2005, 32(4): 614-619.
[26] 张荣铣, 刘晓忠, 宣亚南, 等. 小麦叶片展开后光合碳同化能力—叶源量的估算[J]. 中国农业科学, 1997, 30(1): 84-91. Zhang R X, Liu X Z, Xuan Y Netal. The capacity of photosynthetic assimilation of carbon after leaf expansion in wheat-an estimation of leaf source capacity(LSC)[J]. Sci. Agric. Sin., 1997, 30(1): 84-91.
[27] 江力, 曹树青, 张荣铣, 等. 两个品种烟草叶片发育过程中几种光合参数变化的比较[J]. 植物生理学通讯, 2006, 42(6): 1050-1054. Jiang L, Cao S Q, Zhang R Xetal. Comparison on changes of several photosynthetic parameters in leaf developing process between two tobacco cultivars[J]. Plant Physiol. J., 2006, 42(6): 1050-1054.
Photosyntheticcharacteristicsoffullexpansionleavesofflue-curedtobaccoindifferentelevations
JIANG Hou-long1, WANG Rui2*, JIA Feng3, ZHANG Jun4, ZHANG Zheng-yang5
(1ChongqingTobaccoScienceResearchInstitute,Chongqing400715,China; 2EnshiPrefectureCompanyofHubeiProvincialTobaccoCorporation,Enshi,Hubei445000,China; 3CollegeofBioengineering,HenanUniversityofTechnology,Zhengzhou450001, 4HenanUniversityofScienceandTechnology,Luoyang,Henan471003,China; 5JiangxiProvincialTobaccoCorporation,Nanchang330025,China)
2013-02-25接受日期2013-06-28
湖北省科技厅科技攻关项目“清江流域优质烟叶基地开发与工业利用研究”(2006AA201C73)资助。
江厚龙(1980—)男,河南信阳人,农艺师,博士,从事烟草栽培与生理生态研究。E-mail: jhl513@163.com。 * 通信作者 E-mail: wangrui2999@126.com。
S572
A
1008-505X(2013)06-1483-11
猜你喜欢
中国甜菜糖业(2022年4期)2023-01-21
散文诗(2021年22期)2022-01-12
农业与技术(2020年12期)2020-07-04
少儿科技(2019年9期)2019-09-10
环球时报(2019-05-23)2019-05-23
农学学报(2019年1期)2019-02-21
新课程·下旬(2017年7期)2017-08-14
郑州大学学报(工学版)(2014年6期)2014-03-01
中国烟草学报(2012年2期)2012-04-09
中学生物学(2008年6期)2008-08-29
