
猪原发性心肌症相关蛋白1基因的多态性分析
2013-09-20 13:24田亚光杨秀芹
东北农业大学学报 2013年12期
田亚光,郑 鹏,黄 贺,杨秀芹
(东北农业大学动物科学技术学院,哈尔滨 150030)
原发性心肌症相关蛋白1(Cardiomyopathy-as⁃sociated protein 1,CMYA1)基因最早通过mRNA差异显示技术在鸡胚的房室沟中发现,又命名为XIRP1或Xin基因[1]。利用原位杂交、Northern blot和Western blot杂交技术在mRNA和蛋白水平上研究发现,该基因是骨骼肌特异表达基因,其编码产物位于成年动物心脏闰盘的连接处,可能参与心脏闰盘的形成及维持肌原纤维的完整性[1-2]。CMYA1能够结合肌动蛋白丝,并组织微丝成网状结构[3]。小鼠Xinα基因敲除后表现为心脏肥大,超显微结构显示心脏闰盘处具有明显的分裂及肌纤维肌丝紊乱[4]。进一步研究发现Xin与细胞内信号传导和细胞骨架重塑有关[3、5],参与BMP2-Nkx2.5-MEF2C通路来调控心脏形态变化[2-4]。Hawke等研究发现,作为一种肌动蛋白,Xin转录量在肌肉受损后的12 h急剧增加(大于16倍)[6]。因此,CMYA1与心肌和骨骼肌生长发育密切相关。
不同物种CMYA1蛋白都含有5个高度保守的结构域,分别为富含脯氨酸的区域、16个氨基酸的重复序列(又叫Xin重复序列)、DNA结合结构域、SH3结构基序和核定位信号。猪CMYA1基因由2个外显子组成,编码区全长5 420 bp,预期编码1 839个氨基酸残基的多肽链,定位在13号染色体上[7]。通过比对猪QTL数据库,发现CMYA1可能与猪平均背膘厚、第一肋骨处背膘厚、平均日增重、腿臀重以及屠宰率等性状有关。许晓玲等研究认为猪CMYA1基因与背膘厚存在显著相关[8],但孙泰雷等在13/17罗伯逊异位杂合子猪中的研究却未得到相同结果[9]。本研究以猪CMYA1基因编码区存在的3个错义突变位点c.1394A>G(p.His465 Arg)、c.1751A>G(p.Asp582Gly)和 c.3290C>A(p.Thr1097Asp)为研究对象,利用PCR-RFLP方法研究不同基因型在北京黑猪、丹系长白猪、大白猪和法系长白猪4个品种中的分布情况,为进一步揭示CMYA1基因与生长和胴体性状间关系提供依据。
1 材料与方法
1.1 组织样和DNA提取
采集191头猪的耳组织样,其中北京黑猪(39头)和法系长白猪(31头)采自黑龙江省农业科学院猪场,大白猪(53头)和丹系长白猪(68头)来自哈尔滨香坊猪场。利用酚-氯仿法提取基因组DNA[9]。
1.2 引物的设计与合成
根据GenBank提供的猪CMYA1基因mRNA序列(No.EF173403),针对3个待研究的变异位点设计引物,为后续利用PCR-RFLP方法进行变异位点检测和基因分型提供条件。引物序列及扩增条件见表1。
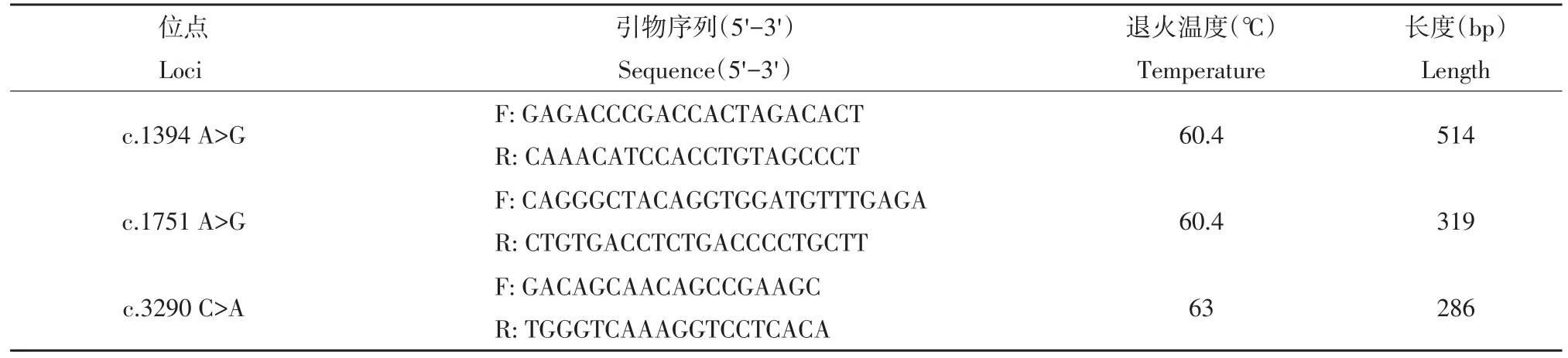
表1 引物序列及扩增条件Table1 Primer sequences and amplification conditions
1.3 基因型检测
PCR扩增条件:15 μL的反应体系中包括10×PCR buffer 1.5 μL、dNTPs mix 0.6 μL、上下游引物(10 μmol·L-1)各0.6 μL、rTaq DNA聚合酶 1 U、模板DNA 50 ng。反应条件为95℃预变性5 min后,95℃变性30 s,60.4℃/63℃退火30 s,72℃延伸45 s,循环33次,72℃终延伸5 min。
对不同突变位点的PCR产物分别用NcoI(c.1394A>G)、HaeⅢ(c.1751A>G)和RsaI(c.3290 C>A)进行酶切。
酶切体系、反应时间及产物检测参照文献[10]进行,酶切温度按照说明书要求设置。
1.4 统计分析
使用SPSS软件进行χ2独立性和适合性检验,用PIC-Calc 0.6软件进行多态信息含量(Polymor⁃phism information content,PIC)计算。
2 结果与分析
2.1 PCR-RFLP结果统计
针对3个位点设计的引物分别成功扩增出特异性很好的PCR产物,酶切后都得到多态性条带。其中,c.1394A>G位点的PCR产物酶切后得到514 bp、514 bp/347 bp/167 bp和347 bp/167 bp 3种带型,分别命名为AA、AB和BB基因型;c.1751A>G位点的PCR产物酶切后得到319 bp、319 bp/212 bp/107 bp和212 bp/107 bp 3种带型,分别命名为AA、AB和BB基因型;c.3290C>A位点的PCR产物酶切后得到286 bp、286 bp/132 bp/154 bp和132 bp/154 bp 3种带型,分别命名为AA、AB和BB基因型。各位点在不同品种中的检测结果见表2。
2.2 统计参数计算
根据PCR-RFLP统计结果,计算得到3个多态位点的纯合度、杂合度、多态信息含量等统计参数值,结果发现c.1394A>G和c.1751A>G都属于中度多态位点(0.24<PIC<0.5)(见表3)。
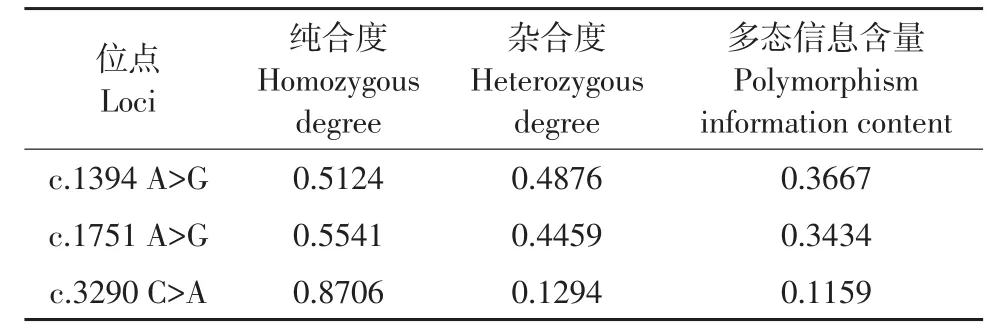
表3 猪CMYA1基因3个位点群体遗传学分析Table3 Statistic values of 3 polymorphic loci of pig CMYA1 gene
2.3 适合性检验
χ2适合性检验表明,在c.1394A>G多态位点,所检测的4个群体中基因型的分布都符合Hardy-Weinberg平衡定律;在c.1751 A>G多态位点,只有北京黑猪是Hardy-Weinberg平衡群体;在c.3290 C>A多态位点,基因型在北京黑猪中的分布不符合Hardy-Weinberg平衡定律(见表4)。

表4 哈代-温伯格平衡检验结果Table4 Result of Hardy-Weinberg equilibrium test
2.4 独立性检验
χ2独立性检验表明在各多态位点,基因型在品种间的分布都存在显著差异(P<0.05),各基因型在品种间分布情况的多重比较结果见表5。

表5 不同位点在各品种(系)间基因型分布的多重比较结果Table5 Multiple comparisons of individual genotypes of 3 polymorphic sites among breeds(lines)
3 讨论与结论
我国是猪肉生产和消费大国,猪肉消费量占全球一半。上世纪九十年代以前,猪遗传育种以改良生长繁育为主要目标;现阶段猪的育种目标不但追求良好的生长繁育性状,还要提高猪的抗病能力。
许晓玲等在获得杜洛克和二花脸猪背最长肌差异表达EST基础上,克隆获得CMYA1基因[8]。刘榜等在伯克夏与约克夏杂交群体中确定CMYA1基因与猪的背膘厚存在显著相关,是一个与背膘厚相关的QTL[7]。另外,CMYA1蛋白与心肌发育和形态变化有关,可以促进闰盘器的成熟和稳定,保证心脏正常功能[4];在构建心肌肥厚和高血压动物模型中,Xin蛋白显著上调表达[11];Julia等[12]在人类心脏组织中检测XinC蛋白表达时,发现其仅在心肌肥厚患者的样本中存在;这些都说明该基因与心血管疾病发生存在着相关性。因此,对猪CMYA1基因的研究,不但在生长和胴体性状上有潜在应用价值,也有可能与猪抗病育种相联系。
CMYA1基因在哺乳动物中高度保守,人CMYA1基因编码1 843个氨基酸,牛CMYA1基因编码1 820个氨基酸。猪CMYA1基因的核苷酸序列与人和牛相应序列相似性都为87%,氨基酸水平上的相似性则分别为83%和78%。本研究所检测的3个多态位点中,c.1394A>G(p.His465Arg)和c.1751 A>G(p.Asp582Gly)位点紧邻16个氨基酸的重复序列(见图1)。

图1 2个多态位点在氨基酸序列上的位置及同源性比对Fig.1 Positions and homology analysis of the 2 out of 3 polymorphic sites
16个氨基酸重复序列是肌动蛋白的结合结构域[6、13],并参与组建细胞骨架的微丝网络[14]。此外,c.1751 A>G(p.Asp582Gly) 在人、猪、狗、牛、猕猴、小鼠等物种中高度保守(见图1),这两个位点变异可能会对CMYA1功能产生一定影响。
c.1394A>G(p.His465Arg)和 c.1751A>G(p.Asp582Gly)2个点突变在大白、丹系长白、法系长白和北京黑猪中的分布存在极显著差异,并且发生频率较高(>5%);结合同源性和结构功能域分析结果,该突变具有作为遗传标记的潜在可能性。
[1]Wang D Z,Hu X,Lin J L,et al.Differential display of mRNAs from the atrioventricular region of developing chicken hearts at stages 15 and 21[J].Front Biosci,1996(1):1-15.
[2]Wang D Z,Reiter R S,Lin J L,et al.Requirement of a novel gene,Xin,in cardiac morphogenesis[J].Development,1999,126(6):1281-1294.
[3]Pacholsky D,Vakeel P,Himmel M,et al.Xin repeats define a novel actin-binding motif[J].J Cell Sci,2004,117(22):5257-5268.
[4]Lin J J C,Gustafson-Wagner E A,Sinn H W,et al.Structure,ex⁃pression,and function of a novel intercalated disc protein,Xin[J].J Med Sci,2005,25(5):215-222.
[5]van der Ven P F M,Ehler E,Vakeel P,et al.Unusual splicing events result in distinct Xin isoforms that associate differentially with filamin c and Mena/VASP[J].Exp Cell Res,2006,312(11):2154-2167.
[6]Hawke T J,Atkinson D J,Kanatous S B,et al.Xin,an actin bind⁃ing protein,is expressed within muscle satellite cells and newly regenerated skeletal muscle fibers[J].Am J Physiol Cell Physiol,2007,293(9):1636-1644.
[7]刘榜,许晓玲,李奎,等.一种猪背膘厚基因CMYA1的克隆及其应用[P].中国,200510019912.4.2005-11-29.
[8]Xu X L,Xu X W,Pan P W,et al.Porcine skeletal muscle differen⁃tially expressed gene CMYA1:isolation,characterization,map⁃ping,expression and association analysis with carcass traits[J].Anim Genet,2009,40(3):255-261.
[9]魏巍,王希彪,黄宣凯,等.民猪群体氟烷基因频率检测及序列多态性分析[J].东北农业大学学报,2011,42(9):31-36.
[10]杨秀芹,刘娣,李景芬,等.猪myostatin基因5'调控区的酶切多态性分析[J].东北农业大学学报,2002,33(3):209-212.
[11]Wang Q,Lin J L C,Wu K H,et al.Xin proteins and intercalated disc maturation,signaling and diseases[J].Front Biosci,2012,17:2566-2593.
[12]Otten J,van der Ven P F M,Vakeel P,et al.Complete loss of mu⁃rine Xin results in a mild cardiac phenotype with altered distribu⁃tion of intercalated discs[J].Cardiovasc Res,2010,85(4):739-750.
[13]Cherepanova O,Orlova A,Galkin V E,et al.Xin-repeats and nebulin-like repeats bind to F-actin in a similar manner[J].Mol.Biol,2006,356(3):714-723.
[14]Pacholsky D,Vakeel P,Himmel M,et al.Xin repeats define a novel actin-binding motif[J].J Cell Sci,2004,117(22):5257-5268.
猜你喜欢
新世纪智能(高一语文)(2020年12期)2020-06-01
中国洗涤用品工业(2019年4期)2019-05-11
中成药(2018年1期)2018-02-02
中成药(2017年3期)2017-05-17
科技视界(2016年27期)2017-03-14
中国医药导报(2015年27期)2015-02-28
现代检验医学杂志(2015年6期)2015-02-06
郑州大学学报(理学版)(2014年3期)2014-03-01
作物研究(2014年6期)2014-03-01
中国神经精神疾病杂志(2013年1期)2013-03-11
