
ADD1基因多态性位点与猪肌内脂肪含量、嫩度及背膘厚的关联性研究
2013-09-12 07:39李江凌陈晓晖龚建军赵素君谢廖党金何志平吕学斌
中国猪业 2013年6期
李江凌 刘 锐 陈晓晖 龚建军 赵素君谢 晶 廖党金 何志平 高 荣 吕学斌
(1四川省畜牧科学研究院,四川成都 610066;2四川大学,四川成都 610000)
ADD1基因多态性位点与猪肌内脂肪含量、嫩度及背膘厚的关联性研究
李江凌1刘 锐1陈晓晖1龚建军1赵素君1谢 晶1廖党金1何志平1高 荣2吕学斌1
(1四川省畜牧科学研究院,四川成都 610066;2四川大学,四川成都 610000)
将82头藏猪和84头其他品种猪屠宰后测定背膘厚,采集背部最长肌测定肌内脂肪含量和嫩度。采集血样提取DNA,利用PCR-RFLP分析ADD1基因的基因型,发现藏猪和其他猪种中均能检测到三种基因型AA、AB和BB。三种基因型的肌内脂肪含量和嫩度存在差异(P<0.05),而背膘厚不存在差异(P>0.05)。ADD1基因在藏猪和其他猪种中存在多态性位点,可以作为肌内脂肪选育的标记辅助选择位点。
脂肪细胞定向和分化决定因子1(ADD1);PCR-RFLP;肌内脂肪;藏猪
在过去的猪新品种选育中曾经把提高瘦肉率、提高生长速率作为主要目标,不过随着瘦肉率提高、生长速度提高,导致肌肉脂肪含量下降,猪肉品质下降。而我国的一些地方猪种如藏猪等具有较高的肌内脂肪含量,猪肉品质较好,瘦肉率却普遍较低。因此,目前与猪肉品质密切相关的肌内脂肪含量成为猪育种工作中关注的一个重点性状。
肌内脂肪含量是一个遗传力较高的性状,但是在活体状态时无法直接测定,常规的育种方法不能进行有效的选育。筛选与肌内脂肪含量相关的候选基因,进行标记辅助选择是进行肌内脂肪含量有效选育方法。近年来对与肌内脂肪含量相关基因的研究较多,但研究结果不尽一致。脂肪细胞定向和分化因子1(ADD1) 基因又称固醇调节元件结合蛋白 -1(sterol-regulatoryelement-binding protein-1SREBP1)基因,编码脂肪细胞定向和分化因子1[1]。ADD1基因是脂肪细胞分化过程中一种重要的核转录因子,通过氧化物酶体增殖蛋白激活受体(PPARγ) 和CCAAT元件增强子结合蛋白(C/EBPs) 共同控制脂肪细胞的分化过程[2],同时还直接参与调节与脂肪酸、甘油三酯和葡萄糖代谢相关的酶基因的表达[3]。已有的研究显示,在正常的非脂肪组织,脂肪含量的稳定依赖于ADD1对脂肪合成的调控[4]。如果ADD1基因超水平表达,就可以使如肌肉、肝脏等非脂肪组织内脂肪细胞沉积。Kakuma(2000)等[5]发现ADD1基因高表达与肌肉、肝脏等非脂肪中脂肪沉积有着确定的关系。Soazig(2002) 等[6]的研究结果表明ADD1在脂肪细胞分化过程中发挥重要的作用。本试验的目的是对ADD1基因的extron2进行多态性分析,研究82头藏猪群体和84头其他猪群体中基因的多态性与肌内脂肪含量、嫩度以及背膘厚之间的关系,为寻找合适的遗传标记提供基础资料。
1 材料与方法
1.1 试验动物
试验动物为四川省畜牧科学研究院简阳试验基地82头藏猪和84头其他品种猪,采集血样后屠宰,取背部最长肌3c m左右,一部分液氮中冷冻,-80℃保存,一部分-20℃保存。
1.2 肉质性状测定
在右酮体胸腰结合处用游标卡尺测定背膘厚;采用索氏抽提法测定肌内脂肪含量。采用TAXTPlus嫩度仪测定嫩度。
1.3 基因多态性分析
采用TIANamp Genomic DNA试剂盒提取猪血样D N A(天根生化科技有限公司)。
ADD1基因引物参照Gen Bank中猪的mRNA序列(序列号:DQ519396.1)设计,引物序列为:上游引物:5′-G C G A C G G T G C C T C T G G T A G T C A T-3′;下游引物:5′-C G C A A G A C G G C G G A T T T A C G G C A-3′。
ADD1基因PCR反应体系:2×Master Mix12.5μL, cDNA模 板0.5μL, Primer1(10μM)1μL,Primer 2(10μM)1μL, d d H2O 加 至25μL。PCR反应条件:94℃4分钟;94℃30秒、55℃30秒、72℃30秒,28个循环;72℃延伸5分钟。
PCR产物进行RFLP,用限制性内切酶HinfⅠ酶切后,3%琼脂糖凝胶电泳检测酶切结果,凝胶成像系统观察并记录。
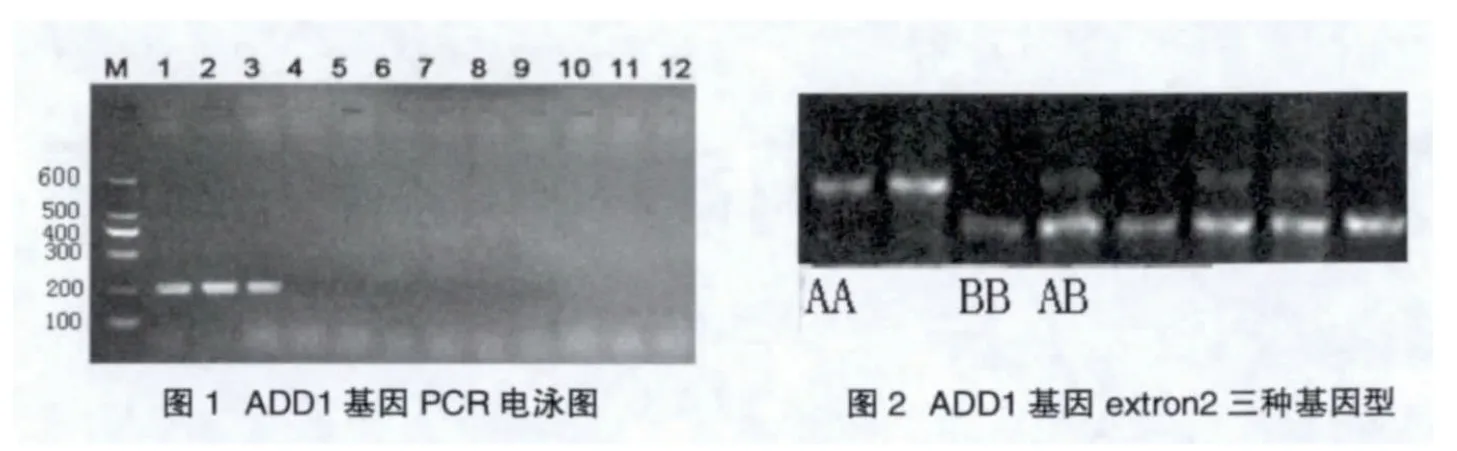

表1 ADD1基因基因型及基因频率分布
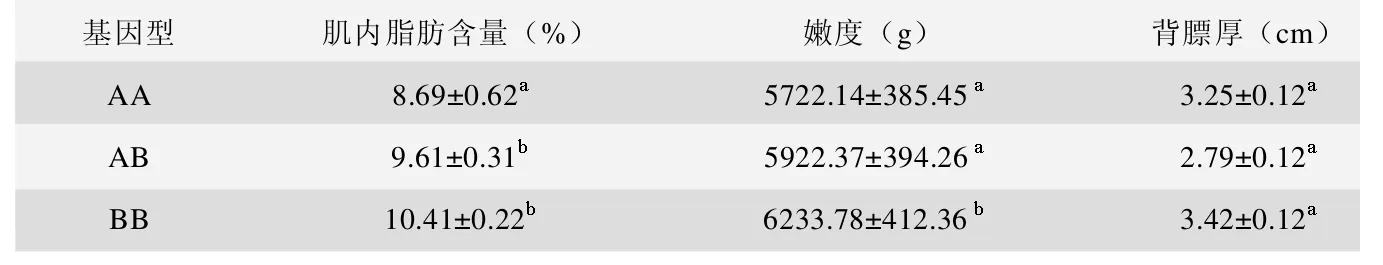
表2 ADD1基因不同基因型的肉质性状差异
1.4 数据分析
ADD1各基因型间肌内脂肪含量、嫩度和背膘厚差异采用S P S S 11.0进行单因子方差分析。
2 结果与分析
PCR扩增产物长度为220b p(图1)。对PCR产物进行RFLP分析,发现该基因产生了3种基因型,纯合子为1条泳带,有两种分别命名为AA,BB,杂合子有两条泳带命名为AB(图2)。三种基因型在本研究所用的藏猪和其他猪种中均能检测到(表1),说明在藏猪群和其他猪群中,ADD1基因存在多态性位点。在藏猪群体中B B基因型频率最高,A B型次之,A A型最少,等位基因B出现频率高于等位基因A出现频率;而在其他猪群中,三种基因型的频率相当,等位基因A和B的出现频率相当。这一结果是否与藏猪的肌内脂肪含量高等优质性状相关还有待于进一步研究。
进一步将所有的试验组猪按基因型进行分类,比较了不同基因型间肉质性状的差异(表2),发现基因型B B和基因型A B肌内脂肪含量显著高于基因型A A(P<0.05),基因型B B嫩度显著高于其他两种基因型(P<0.05),而三种基因型的背膘厚无显著差异(P>0.05)。ADD1基因不同基因型间背最长肌内脂肪含量、嫩度存在差异,而不同基因型的背膘厚却没有显著差异,提示在该基因上发现的这个标记可能可以作为猪肌内脂肪选育的辅助标记。
[1] Edwards P A, Tabor D, Kast H R,et al. Regulation of gene expression by SREBP and SCAP. Biochimica et Biophysica Acta,2000,1529 (1-3):103-113.
[2] Fleischmann M, Iynedjian P B.Regulation of sterol regulatory-element binding protein 1 gene expression in liver:role of insulin and protein kinase B/ cAkt.Biochem J,2000,349:13-17.
[3] Kotzka J, Wieland D, Koponen A, et al. ADD1/ SREBP-1c mediates insulin -induced gene expression linked to the MAP kinase pathway [J] . Biochem Biophys Res Commun, 1998, 249 (2):375-379.
[4] Foretz M, Guichand P, Ferre P, et al.SREBP-1c is a major mrfiator of insulin action on the hepatic expression of gluckinase and lipogenesis related genes. Proc Natl Acad Sci USA,1999,(96):12737-12742
[5] Kakuma T,Lee Y, Higa M, et al.Leptin troglitazone, and the expressing of sterol regulatory element binding proteins in liver and pancreatic islets. Proc Natl Acad Sci USA,2000,(97):8536-8541
[6] SoazigL L, IsablleL,Christian T, et al.Insulin and sterol-regulatory elemenet-binding protein-1c (SREBP-1c)reregulation of gene expression in 3T3-L1 adipocytes. J Biol Chem, 2002, 277:35625-35634
S813.3
A
1673-4645(2013)06-0044-02
2013-06-07
四川省基本科研业务费专项资金项目SASA 2008B 01;四川省繁育专项S A S A 2009Y Z 001
李江凌(1972-),研究员,主要从事动物遗传育种研究,e-ma i l:yujiang 1465@126.com
吕学斌(1965-),研究员,主要从事猪遗传育种研究
猜你喜欢
猪业科学(2022年10期)2022-11-03
世界科学技术-中医药现代化(2022年3期)2022-08-22
昆明医科大学学报(2022年2期)2022-03-29
猪业科学(2022年1期)2022-03-24
猪业科学(2021年6期)2021-08-12
昆明医科大学学报(2021年3期)2021-07-22
猪业科学(2021年3期)2021-05-21
肉类研究(2017年10期)2017-11-20
肉类研究(2016年6期)2016-07-22
肉类研究(2016年4期)2016-05-17
