
不同呈色早熟桃叶片夜间呼吸速率的影响因素
2013-08-22 03:25:22彭丽丽姜卫兵王明玉张斌斌马瑞娟
江苏农业学报 2013年5期
彭丽丽,姜卫兵,韩 健,王明玉,张斌斌,马瑞娟
(1.南京农业大学园艺学院,江苏 南京 210095;2.江苏省农业科学院园艺研究所,江苏 南京 210014)
植物夜间呼吸作用是影响植物生长发育的重要因素,它的代谢过程复杂,受自身遗传因素和环境因子的双重影响。目前,关于植物叶片夜间呼吸作用的研究报道主要集中在设施蔬菜上[1-2],尚未见自然条件下早熟桃夜间呼吸作用生理特性及其与环境因子关系的报道。本研究拟选用早熟夏秋红叶性状典型的早美和绿叶品种锡蜜,尝试用灰色系统理论来初步探讨不同生长发育时期不同呈色早熟桃叶片夜间呼吸作用与环境因子及其碳水化合物的关系。
1 材料与方法
1.1 供试材料
试验于2012年3~8月在江苏省农业科学院国家果树种质南京桃资源圃中进行。试验材料为生长发育基本一致的7年早熟桃(夏秋红叶性状典型的早美和绿叶锡蜜)的成年树,砧木为毛桃,株行距5 m×3 m,树势中庸,干周42~44 cm。南北成行种植,相互不遮挡。试验地以棕壤土为主,肥力中等。
1.2 试验方法
为避免果实库的影响,在3月31日至4月2日,对试验材料进行完全抹除花苞处理。在每株树外围的西南、南向选择长势基本一致、无病虫害的长果枝,以枝梢中部成熟的功能叶为测定对象。分别在6、7、8月下旬的晴朗夜晚21∶00,测定主要环境因子、叶片呼吸速率(Rn)、叶片总可溶性糖和淀粉含量等指标,3次重复。
1.2.1 环境因子的测定 利用浙江托普仪器有公司生产的TNHY-9型手持式农业环境监测仪测定晚上21∶00时大气 CO2(Ca)、大气温度(Ta)和湿度(H),3 次重复。
1.2.2 夜间呼吸速率的测定 自然条件下,选取完好的功能叶,采用CIRAS-1型光合测定系统(PPSystem,英国)开放式气路,测定21∶00时叶片的Pn、Ci、Gs、Tr,Pn绝对值相当于夜间呼吸值(Rn),水分利用效率WUE=Rn/Tr,3次重复。
1.2.3 可溶性糖和淀粉的测定 测定呼吸作用的同时,采集叶片,带回实验室保存于4℃冰箱,次日采用蒽酮法测定可溶性糖和淀粉含量[3]。
1.3 数据统计与分析
利用Excel和SPSS软件进行数据统计与差异性分析。灰色关联度分析法参照戴美松等[4]的方法。
2 结果
2.1 试验环境因子的测定
试验期间夜晚21∶00外界环境因子温度(Ta)、湿度(H)、二氧化碳浓度(Ca)如表1所示。7月下旬温度显著大于6月和8月下旬,后二者差异不显著;湿度相反;Ca大小为:6月>7月>8月。相关性分析结果表明,Ca与Ta,Ca与H相关性不显著(P>0.05);Ta与 H极显著负相关,相关系数为-0.956(P <0.01)。
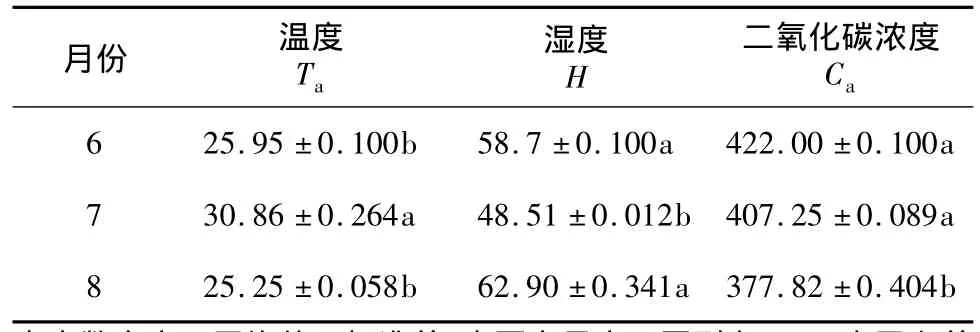
表1 试验期间环境因子Table 1 Environment factors during the experiment
2.2 不同生长发育时期早熟桃叶片Rn、WUE、Ci、Ti及Gs的变化
不同生长发育时期早美、锡蜜叶片夜间呼吸速率(Rn)、水分利用效率(WUE)、胞间CO2浓度(Ci)、气孔导度(Gs)及叶表面温度(Ti)的变化如表2所示。6月和7月下旬,早美Rn、WUE及Ci显著大于锡蜜(P<0.05),8月下旬二者相反;早美Ti在6月和7月下旬与锡蜜差异不显著(P>0.05);早美Gs在6月和8月下旬大于锡蜜,6月下旬差异达到显著水平(P<0.05),7月下旬二者大小相反,且达到显著水平(P<0.05)。
早美 Rn、WUE、Ci、Ti大小均为:7 月 >8 月 > 6月;Gs为:6月>8月>7月。锡蜜Rn为:6月<7月<8月;WUE、Ci均为:8月>6月 >7月;Ti分别为:7月>8月>6月;Gs为7月>6月>8月。
2.3 不同生长发育时期早熟桃叶片总可溶性糖和淀粉含量的变化
不同生长发育时期早美、锡蜜叶片夜间总可溶性糖和淀粉含量如表3所示。6月和7月下旬,早美总可溶性糖含量均大于锡蜜,8月相反。6月和8月下旬,早美淀粉含量显著小于锡蜜(P<0.05),7月相反,但未达显著水平(P>0.05)。
测定期内,早美叶片总可溶性糖含量大小为:7月>8月>6月,淀粉相反。而锡蜜总可溶性糖、淀粉含量大小均为:6月<7月<8月。此外,6月下旬至8月下旬,早美淀粉含量增幅较锡蜜小,分别为45.78%、49.60%。

表2 不同生长发育时期早熟桃叶片夜晚Rn、WUE、Ci、Tl及GS的变化Table 2 Changes of Rn,WUE,Ci,Tland GSof early-maturing peach leaves during growth and development at night

表3 不同生长发育时期早熟桃叶片夜晚碳水化合物含量的变化Table 3 Changes of carbohydrates of early-maturing peach leaves during growth and development at night
2.4 不同生长发育时期早熟桃叶片Rn与影响因素的关联分析
2.4.1 Rn与环境因子 不同生长发育时期早美、锡蜜叶片夜间呼吸速率(Rn)与环境CO2浓度(Ca)、温度(Ta)、湿度(H)及叶表面温度(Ti)的关联度及其排序见表4。关联度分析结果显示,3个测定期影响锡蜜Rn的主要环境因子是Ti,6月和8月下旬次要环境因子是 Ca,7月下旬则是Ta,这可能是夜间21∶00时温度达到30.86℃,细胞呼吸作用所需呼吸酶活性仍较高。6、7、8月下旬影响早美Rn的主导环境因子分别为Ca、H、Ta。

表4 不同生长发育时期早熟桃夜间呼吸速率与环境因子的关联度Table 4 Correlation degree between Rnand environmental factors of early-maturing peach leaves during growth and development at night
2.4.2 Rn与WUE、Ci及Gs不同生长发育时期早美、锡蜜叶片Rn与WUE、Ci及Gs的关联度及排序见表5。6月和7月下旬,锡蜜Rn与WUE、Ci及Gs的关联度排序均为 WUE、Ci、Gs,8月下旬排序为:WUE、Gs、Ci;6月和7月下旬主导早美Rn的是Ci,8月主导其 Rn的是 WUE,Ci次之。
2.4.3 Rn与碳水化合物 不同生长发育时期早美、锡蜜叶片夜间呼吸速率(Rn)与总可溶性糖和淀粉的关联度及排序见表6。灰色关联分析结果表明,早美在6月和7月下旬Rn与淀粉含量关联度大于Rn与可溶性糖含量,8月相反;锡蜜在6月下旬Rn与淀粉含量的关联度小于Rn与可溶性糖含量,7月和8月下旬反之。

表5 不同生长发育时期早熟桃夜间呼吸速率与WUE、Ci及Gs的关联度Table 5 Correlation degree between Rnand WUE,Ciand Gsof early-maturing peach leaves during growth and development at night

表6 不同生长发育时期早熟桃夜间呼吸速率与碳水化合物含量的关联度Table 6 Correlation degree between Rnand carbohydrate content of early-maturing peach leaves during growth and development at night
3 讨论
植物呼吸作用实际上是糖类物质降解氧化的过程,主要涉及糖酵解、戊糖磷酸途径和三羧酸循环3种代谢途径,其中糖酵解第1阶段是淀粉或己糖的活化[5]。据研究,可溶性糖通过糖酵解途径参与花色素苷的合成,其中,糖代谢涉及己糖代谢,己糖激酶和果糖激酶分别催化葡萄糖和果糖的磷酸化,是己糖进入糖酵解的第一步反应,也是不可逆反应[6-7],糖通过己糖激酶磷酸化相关的信号转导途径调节花色素苷合成[8]。综合锡蜜、早美叶片夜间呼吸作用表明,6、7月下旬早美叶片夜间呼吸作用糖酵解第1阶段是淀粉的活化,生成己糖,参与代谢;7月下旬夜间温度达到30.86℃,植株代谢旺盛,更多淀粉通过呼吸作用降解,参与植株代谢。同时,本研究结果显示,8月下旬早美叶片完成转色初期[9],其呼吸作用糖酵解第1阶段是己糖的活化,锡蜜则是淀粉;这是因为早美叶片转色完成初期对己糖需求量降低,此外,此期温度相对较低,己糖含量降低,从而使得锡蜜呼吸消耗淀粉。
植物呼吸作用一般最适温度是25~35℃,其呼吸速率随着外界CO2浓度的上升而下降[5]。汪杏芬等[10]认为,较低温度下,CO2浓度倍增对植物暗呼吸没有显著影响;只有在较高温度(30℃、35℃)下多数植物的暗呼吸显著增强。本研究结果表明,6月下旬夜间温度较高(25.95℃),较高的 CO2浓度(422 μl/L)和胞间CO2浓度主导早美叶片呼吸速率。7月下旬,主导早美叶片呼吸速率的是环境湿度和胞间CO2浓度,这可能是因为白天光照强度、温度达到周年最高,导致环境湿度较低所致;8月下旬环境温度和叶片水分利用效率是影响早美夜间呼吸速率的主要因素。
植物夜间呼吸速率不仅与温度、CO2浓度等环境因子密切相关,也与植物特性有关[11-12]。植物叶片处于幼叶时呼吸较快,成长后即开始下降,到衰老时由于氧化磷酸化开始解偶联,能量传递体系破坏,P/O比明显下降,导致呼吸上升[5]。本研究结果显示,早美叶片呼吸速率先增后降,锡蜜则缓慢增加,表明花色素苷能阻止氧化磷酸化。
综上,早熟桃叶片夜间呼吸速率除了受环境因子的综合影响外,也与叶片色彩的表达代谢密切相关。
[1]李 梅,须 晖,李天来,等.不同夜温对番茄叶片呼吸及光合启动时间的影响[J].石河子大学学报:自然科学版,2006,24(2):202-204.
[2]须 晖,孙 博,李 梅,等.不同夜温对茄子苗期单叶呼吸速率和光合启动时间的影响[J].植物生理学通讯,2009,45(5):429-432.
[3]王学奎.植物生理生化实验原理和技术[M].北京:高等教育出版社,2006:202-204.
[4]戴美松,姜卫兵,庄 猛.大棚栽培条件下葡萄叶片净光合速率与其他生理、生态参数的灰色关联分析[J].果树学报,2005,22(5):474-478.
[5]潘瑞炽.植物生理学[M].6版.北京:高等教育出版社,2008.
[6]PETREIKOV M,DAI N,GRANOT D,et al.Characterization of native and yeast-expressed tomato fruit fructokinase enzymes[J].Phytochemistry,2001,58:841-847.
[7]陈俊伟,陈子敏,钱皆兵,等.杨梅果实发育进程中的碳水化合物代谢[J].植物生理与分子生物学学报,2006,32(4):438-444.
[8]NETA I,SHOSEYOV O,WEISSD.Sugars enhance the expression of gibberellin 2 induced genes in developing petunia flowers[J].Physiol Plant,2000,109(2):196-202.
[9]谢智华,姜卫兵,韩 健,等.早熟桃夏季红叶现象的生理机制研究[J].园艺学报,2012,39(7):1271-1277.
[10]汪杏芬,白克智,匡廷云.大气CO2浓度倍增对植物暗呼吸的影响[J].植物学报,1997,39(9):849-854.
[11]REICH P B,TJOELKER M G,MACHADO J L,et al.Universal scaling of respiratory metabolism,size and nitrogen in plants[J].Nature,2006,439(26):457-461.
[12]刘福娟,李红梅,吴国辉.植物呼吸作用的影响因素分析[J].农机化研究,2004(3):267.
猜你喜欢
农业技术与装备(2022年6期)2022-08-17 09:46:18
中成药(2018年8期)2018-08-29 01:28:34
水利科技与经济(2017年12期)2017-04-22 03:10:20
西安工程大学学报(2016年6期)2017-01-15 14:08:26
中国粮油学报(2016年1期)2016-02-06 02:16:53
电源技术(2015年11期)2015-08-22 08:50:18
医学研究杂志(2015年9期)2015-07-01 17:28:24
中国当代医药(2015年9期)2015-03-01 02:02:13
河南科技(2014年16期)2014-02-27 14:13:25
郑州大学学报(理学版)(2013年2期)2013-03-11 20:30:25
