
不同鹅品种脂联素基因第2外显子多态性分析
2013-08-23 03:49:34龚道清
江苏农业学报 2013年5期
王 健, 董 飚, 龚道清
(1.江苏农牧科技职业学院,江苏 泰州 225300;2.国家级水禽基因库,江苏 泰州 225300;3.扬州大学动物科学与技术学院,江苏 扬州 225009)
在动物生产中,动物摄入能量首先用于维持,然后多余的才会被用于生产和转化为脂肪等形式储存起来。脂肪组织可以储存能量,还可参与能量代谢和平衡[1]。禽类的脂肪沉积主要集中在皮下脂肪和腹脂,但这些脂肪并不符合现代消费者的需求。影响禽类的脂肪沉积的因素较多,其中遗传和营养是最主要的因素。如何降低禽类的脂肪沉积能力,提高经济效益,降低饲料成本,国内外禽类遗传育种学者参与了动物脂肪沉积的研究,其中功能基因是研究方向之一。脂联素是脂肪组织特异性分泌的一种胶原样细胞因子,人和哺乳动物脂联素基因包含了3个外显子和2个内含子[2-3],禽类脂联素只有2个外显子和1个内含子[4]。脂联素基因最初是在人类上进行了研究,主要认为其与II型糖尿病有显著性关系,然后才逐渐转移到哺乳动物、禽类上进行研究。在鸭、鹅方面的研究主要集中在多态位点的检测、基因克隆、mRNA在不同组织或者时期的表达规律等[5-11]。本文拟寻找出中国部分地方白鹅品种脂联素基因在外显子上可能存在的点突变,检测其在不同鹅品种中分布情况,分析群体遗传参数,为鹅品种鉴定、保存和选育提供参考依据。
1 材料与方法
1.1 试验材料
本试验用太湖鹅、豁眼鹅、四川白鹅、浙东白鹅、皖西白鹅5种白鹅均由国家级水禽基因库所提供,每个品种60只,公母各半。翅静脉采取血样,4%EDTA进行抗凝,采用酚/氯仿法提取基因组DNA,并溶解于水中备用。
1.2 PCR引物的设计
根据鹅脂联素基因DNA序列(GenBank No:EU370686)设计2对引物,扩增第2外显子序列。ADP1上游引物序列:5'-ACTCCTGATCCCAACTCTGT-3'、下游引物序列:5'-GTCGTAGTGGTTCTGCTCGT-3',退火温度为56℃,扩增长度为245 bp;ADP2上游引物序列:5'-ACGAGCAGAACCACTACGAC-3'、下游引物序列:5'-TGAGATGGAGCAAGACCGAG-3',退火温度为55℃,扩增长度为204 bp。
1.3 PCR 扩增
PCR 扩增体系为20.00 μl,包括10 ×PCR buffer(无 Mg2+)2.00 μl、10 mmol/L dNTPs 2.00 μl、25 mmol/L MgCl21.20 μl、10 pmol/L 上下游引物各0.80 μl、5 U/μl Taq 酶 0.12 μl、100 ng/μl DNA 模板 0.80 μl、加 ddH2O 至 25.00 μl。PCR 扩增条件为94℃变性5 min;94℃ 30 s,56/55℃ 30 s,72℃30 s共35个循环;72℃延伸10 min,4℃保温。PCR产物在1.2%琼脂糖凝胶中电泳,结束后用凝胶成像系统检测扩增结果。
1.4 SSCP分型与测序
在5 μl PCR产物中加入5 μl上样缓冲液进行混合,98℃变性10 min,迅速插入冰中,放置5 min,在12%非变性聚丙烯酰胺凝胶中电泳,银染显色、判型。挑选纯合基因型个体的PCR产物切胶回收,连接到PMD18-T载体上转化到大肠杆菌DH5α上,经过鉴定后送上海桑尼生物工程有限公司进行序列测序。
1.5 数据统计分析
运用DNAMAN软件比对寻找出不同基因型间存在的点突变。根据聚丙烯酰胺凝胶染色图统计出各个群体基因型数,进而计算出基因型频率和等位基因频率,用卡方检验分析不同基因型在群体之间差异情况。
2 结果
2.1 PCR 检测结果
将2对引物分别对部分鹅基因组DNA进行PCR扩增,在1.5%琼脂糖电泳上检测,结果显示扩增片段与目的片段大小一致且特异性好,可以直接进行SSCP分析。
2.2 SSCP 检测结果
引物ADP1对不同鹅品种进行PCR扩增后进行SSCP检测,结果显示引物ADP1扩增的片段上存在多态,主要存在3种基因型,分别命名为XX、XY和YY(图1)。引物ADP2对不同鹅品种进行PCR扩增后进行SSCP检测,结果显示引物ADP2扩增的片段上没有多态性。
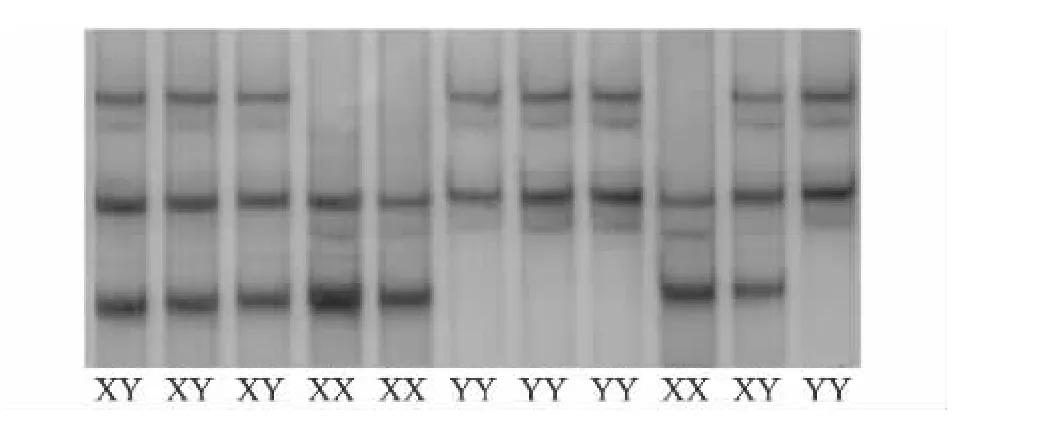
图1 引物ADP1对部分鹅品种PCR扩增产物SSCP分析Fig.1 SSCP analysis of goose breeds PCR products with primer ADP1
2.3 多态性基因片段的克隆与测序
取XX和YY基因型的PCR产物进行片段回收、克隆和测序。运用生物学软件DNAMAN分析测序结果,其中YY型与GenBank中上传的鹅脂联素基因序列一致;XX型与YY型相比,在外显子第351处发生了腺嘌呤(A)→尿嘧啶(G)突变(图2),该突变未引起氨基酸发生变化。

图2 不同基因型间核苷酸序列比较Fig.2 Comparison of nucleotide sequence among different genotypes of goose
2.4 不同鹅品种基因型(基因)频率比较
对不同鹅品种分别进行基因型检测,计算不同鹅品种的基因型频率和基因频率(表1)。太湖鹅以XY基因型较高,YY基因型次之;皖西白鹅、浙东白鹅和四川白鹅均以YY型基因型为主,XY基因型次之,XX基因型比例极小,其中皖西白鹅和浙东白鹅YY型几乎是XY型的2倍;豁眼鹅以XX和XY基因型为主,YY基因型比例相对较低。豁眼鹅以X等位基因为优势基因,其他5个鹅品种均以Y等位基因为优势基因,其中浙东白鹅Y等位基因频率最高,达0.833 3。
由表2可知,豁眼鹅基因型分布与其他鹅群体存在极显著差异;浙东白鹅基因型分布与太湖鹅、四川白鹅存在显著差异;其他鹅品种之间无显著性差异。

表1 不同鹅品种基因型及等位基因频率Table 1 The frequencies of genotypes and alleles among different breeds of goose

表2 不同基因型在鹅品种之间的卡方检验结果Table 2 The χ2detection of genotypes in different breeds of goose
2.5 遗传特性分析
群体遗传分析结果(表3)显示,浙东白鹅纯合度最高,为0.722 2;皖西白鹅次之,为0.670 1;豁眼鹅最低,为0.508 9。除浙东白鹅为低度多态外,其他4个鹅品种为中度多态,其中以豁眼鹅多态信息含量最高,为0.370 5。
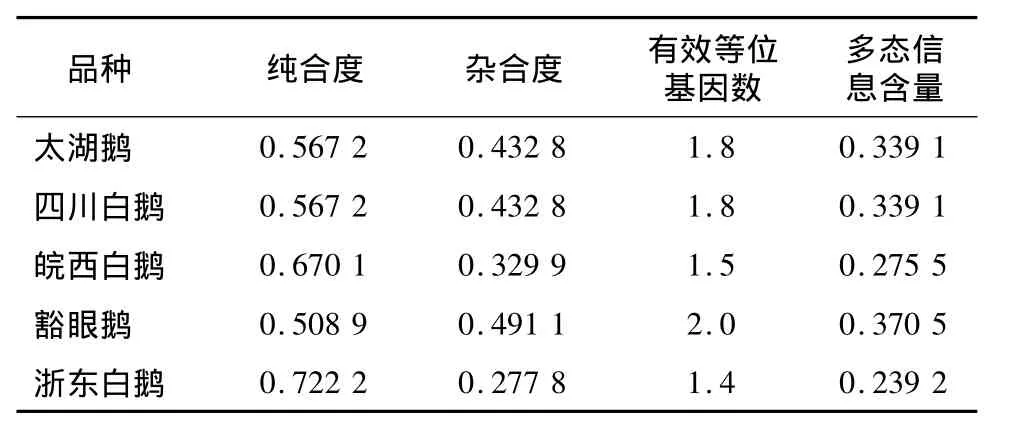
表3 不同鹅品种的遗传参数Table 3 Genetic parameters for different breeds of goose
3 讨论
脂联素基因在禽类群体中均存在,在同一物种或品种,脂联素基因在序列上绝大部分是相同的,只有少数碱基存在差异,这就是所谓的点突变。这些点突变在内含子中的变化影响作用要小于在外显子上,外显子上的点突变可以引起某个氨基酸或者整个氨基酸序列变化,从而影响该序列编码蛋白质的功能。在外显子上点突变主要分为同义突变、错义突变和无义突变。本研究在外显子2上设计了2对引物,首次在鹅脂联素基因外显子上发现了点突变,突变位置为第351处,是腺嘌呤(A)与尿嘧啶(G)之间发生了突变,该突变未引起氨基酸发生变化。与鸭相比,其在外显子2上的点突变数较少[7],这说明鹅在进化过程中比较保守。
在太湖鹅、皖西白鹅、豁眼鹅、四川白鹅和浙东白鹅5个中国地方白鹅群中存在3种基因型,分别为XX型、YY型和XY型。太湖鹅、皖西白鹅、四川白鹅和浙东白鹅主要以XY型、YY型2种形式存在,另外,4种鹅Y等位基因频率均明显高于X等位基因频率。豁眼鹅主要以XX型、XY型2种基因型存在,2个等位基因频率之间的差异相对于其他4个品种来说要小很多。不同等位基因在品种之间分布情况显示,豁眼鹅与其他4个鹅之间存在显著性差异,浙东白鹅与太湖鹅、四川白鹅之间存在显著性差异。豁眼鹅为小型品种,其产蛋率在中国地方品种中最高,主要分布在山东省、辽宁省内;太湖鹅也属于小型品种,主要分布于江苏省和浙江省的太湖流域;浙东白鹅为中型品种,主要分布于浙江省东部。皖西白鹅属于中型品种,主要分布于安徽省西部;四川白鹅属于中型品种,主要分布于四川省。在品种进化过程中,因自然和人类的影响使得不同品种向不同生产性能方向进化,间接导致不同品种之间脂联素基因型频率产生了差异。品种之间的基因型频率差异为品种选育和进行品种杂交提供了素材。在禽类上已经有了一些关于脂联素基因点突变与生产性能之间的关联性分析的报道,刘大林等分析了京海黄鸡脂联素基因第1内含子中的1个点突变与屠体性状的遗传效应,提出脂联素基因可能是调控脂肪性状的主效基因或与主效基因连锁[12]。张依裕等在白羽番鸭内含子中发现了C→T突变,发现CC纯合子的白羽番鸭肉质更好[13]。
本研究以脂联素基因中存在的多态位点分析了5个白鹅群体的遗传参数,以浙东白鹅、皖西白鹅的纯合度较高,豁眼鹅纯合度最低。结合群体遗传信息含量,表明豁眼鹅拥有最丰富的遗传信息,其选择的遗传潜力最大,这与品种形成时间最短也有一定的关系,说明其选择的时间较短;另外,浙东白鹅的遗传信息含量最低。这些结果与董飚等在研究脂联素基因内含子遗传多样性中的分析结果一致[7]。
[1]AILHAUD G.Adipose tissue as a secretary organ:from adipogenesis to the metabolic syndrome [J].Comptes Rendus Biologies,2006,329(8):570-577.
[2]TAKAHASHI M,ARITA Y,YAMAQATA K,et al.Genomic structure and mutations in adipose-specific gene,adiponectin[J].International Journal of Obesity,2000,24(7):861-868.
[3]DAS K,LIN Y,WIDEN E,et al.Chromosomal localization,expression pattern,and promoter analysis of the mouse gene encoding adipocyte-specific secretory protein Acrp30[J].Biochemical and Biophysical Research Communications,2001,280(4):1120-1129.
[4]YUAN J,LIU W,LIU Z L,et al.cDNA cloning,genomic structure,chromosomal mapping and expression analysis of ADIPOQ(adiponectin)in chicken[J].Cytogenetic and Genome Research,2006,112(1-2):148-151.
[5]徐国庆,龚道清,董 飚,等.鹅脂联素基因的克隆、序列分析及组织表达[J].农业生物技术学报,2008,16(6):941-946.
[6]薛茂云,董 飚,顾志良,等.鸭脂联素基因全长cDNA的克隆和原核表达的研究[J].畜牧兽医学报,2010,41(10):1232-1239.
[7]董 飚,龚道清,孟 和,等.鸭脂联素基因单核苷酸多态性检测及群体遗传分析[J].遗传,2007,29(8):995-1000.
[8]杨德全,葛菲菲,刘 健,等.一株鸭源H4N6亚型禽流感病毒A/duck/Shanghai/Y20/2006的全基因组序列测定及遗传演化分析[J].江苏农业学报,2012,28(4):815-822.
[9]钮慧敏,李 银,黄欣梅,等.1株鸭病毒性肝炎病毒的分离与鉴定[J].江苏农业科学,2011,39(6):374-376.
[10]张响英,唐现文,王利刚,等.实时荧光定量RT-PCR检测狮头鹅繁殖周期中PRL mRNA的表达[J].江苏农业科学,2012,40(9):50-52.
[11]洪胜辉,张 军,张 蕊,等.鹅原代肝细胞的简易、高纯分离及培养[J].江苏农业科学,2012,40(4):56-58.
[12]刘大林,俞亚波,魏 岳,等.脂联素基因对京海黄鸡体重及屠体性状的遗传效应[J].扬州大学学报:农业与生命科学版,2009,30(1):31-34.
[13]张依裕,徐 琪,段修军,等.白羽番鸭脂联素基因内含子多态与肉质的关联分析[J].河南农业科学,2010(7):97-103.
猜你喜欢
小猕猴学习画刊(2022年10期)2022-11-01 05:35:26
戏曲研究(2022年2期)2022-10-24 01:53:30
趣味(语文)(2021年12期)2021-04-19 12:16:06
小学生(看图说画)(2020年3期)2020-04-01 11:57:14
工程与建设(2019年2期)2019-09-02 01:33:50
创新作文(小学版)(2016年13期)2016-11-11 05:46:31
胃肠病学(2016年7期)2016-03-13 23:11:56
安徽医科大学学报(2015年9期)2015-12-16 11:09:42
郑州大学学报(医学版)(2015年2期)2015-02-27 14:50:45
中国药业(2014年24期)2014-05-26 09:00:14
