
氨基酸的生物活性及其营养调控功能的研究进展
2013-08-22 06:48王洪荣
动物营养学报 2013年3期
王洪荣 季 昀
(扬州大学动物科学与技术学院,扬州225009)
近年来,越来越多的研究证实了体内某些氨基酸不仅仅作为蛋白质合成的底物原料,它们还能够通过自身及其代谢产物所具有的生物活性对动物机体内许多生命活动产生调节作用,例如调节营养物质代谢(蛋白质代谢、脂代谢、糖代谢等)、为机体提供能够能量、维持机体内环境稳态、合成一氧化氮(NO)、多胺、谷胱甘肽、核酸激素和神经递质,影响神经和内分泌,调控细胞的基因表达和信号转导、免疫、抗氧化、抗应激等功能,这些调节作用最终可影响到动物的生长发育、生产性能以及健康状况。目前研究较多的具有明显生物活性的氨基酸包括精氨酸、谷氨酰胺、亮氨酸、色氨酸、苏氨酸等,本文总结了它们的代谢途径以及营养调节作用的研究进展,为通过氨基酸的营养调控作用来改善动物生产提供参考。
1 精氨酸
1.1 精氨酸的代谢途径
精氨酸是一种半必需氨基酸,在动物体内具有重要的生理、代谢调控和营养作用,其代谢途径见图1。精氨酸通过其自身或其代谢产物NO、多胺、鸟氨酸、脯氨酸等在调节机体代谢和繁殖、促进激素分泌、改善免疫功能、预防心血管疾病和内皮细胞功能紊乱、维持骨骼肌和大脑功能、组织损伤与修复等多个方面均发挥重要的功能[1]。精氨酸是内源性NO的唯一前体,可在一氧化氮合成酶的作用下生成NO,NO在血管扩张过程中作为信号分子物质来促进血管的舒张,提高胰岛素敏感组织的血流量。NO参与调节产能底物代谢(图2),可降低与脂肪合成和糖异生作用相关基因的表达,提高激素敏感脂肪酶磷酸化水平进而促进脂肪分解,此外NO还能够促进线粒体生物合成和氧化磷酸化[2]。精氨酸的代谢产物腐胺、精胺等多胺类物质在维持细胞膜稳定性方面起重要作用;肌酸能够在机体运动时迅速补充能量;鸟氨酸和瓜氨酸是体内尿素循环的重要代谢中间产物。此外,精氨酸也是氨脱毒所必需的物质,保障中枢神经系统的健康[1]。
1.2 精氨酸的营养调控功能
1.2.1 改善动物采食量及畜禽产品品质
精氨酸及其代谢产物可通过影响外周激素的分泌提高畜禽采食量[4]。精氨酸可抑制脂肪细胞增大,降低脂肪沉积。饲粮中添加适宜水平的精氨酸可显著提高环江香猪瘦肉率,显著降低脂肪率和背最长肌中脂肪含量[5]。饲粮中添加0.125%~1.000%的精氨酸可显著提高肉鸭采食量、生产性能、饲料转化效率以及胴体品质,降低皮脂率和肝脂率[6]。Mc Knight等[7]指出了精氨酸减少脂肪合成,提高肌肉含量的可能机理:精氨酸通过改变能量摄取与消耗的平衡状态进而减少脂肪,降低白色脂肪组织的生长,具体的机理可能与精氨酸提高细胞信号分子(NO、多胺、环腺苷酸等)促进线粒体合成和棕色脂肪组织生长有关,也与精氨酸提高能够促进产能底物氧化的基因的表达有关。

图1 精氨酸的主要代谢途径Fig.1 The major metabolic pathway of arginine[3]
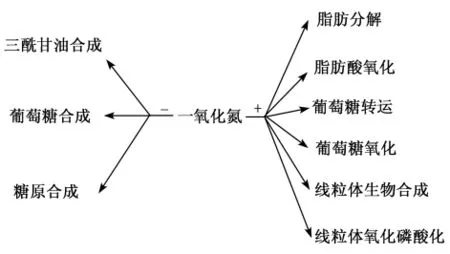
图2 NO对产能底物代谢的调节Fig.2 Regulation of metabolism of substrates of producing energy by NO[1]
1.2.2 调节泌乳功能
由于精氨酸的代谢产物NO具有扩张血管、促进血管生成的作用,因此调控精氨酸-NO途径可以促进乳腺及乳腺内血管的生长,提高营养物质的摄取量,进而改善哺乳动物的泌乳性能[8]。乳腺内精氨酸的摄取量通常情况下高于乳中精氨酸的量,给初产母猪饲粮补充精氨酸或NO前体物可以提高其泌乳性能[8-9]。对于奶牛而言,研究表明真胃灌注精氨酸可显著提高奶牛血中精氨酸、鸟氨酸、尿素含量[10],妊娠后期奶牛颈静脉连续灌注精氨酸能够显著提高血液催乳素、生长激素、胰岛素的含量,进而提高乳产量[11],但也有研究表明当其他必需氨基酸足够的条件下给奶牛真胃灌注无精氨酸的氨基酸混合物对乳产量及乳成分含量几乎没有影响[12],那么,是否精氨酸水平影响奶牛的泌乳性能和乳成分组成有待于进一步研究。细胞分子水平的研究表明,精氨酸水平可调控奶牛乳腺上皮细胞的增殖及酪蛋白基因的表达,可通过调控乳腺上皮细胞内酪氨酸激酶2-信号转导和转录 激 活 子 5(Jak2-Stat5)、雷 帕 霉 素 靶 蛋 白(mTOR)信号转导通路影响酪蛋白基因的转录与翻译(图3)[13-14]。
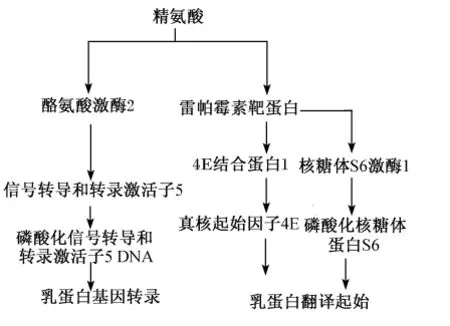
图3 精氨酸对乳蛋白合成调控信号通路的调控Fig.3 Regulation on signaling pathway of milk protein synthesis by arginine[13-14]
1.2.3 改善消化道功能及机体抗氧化和应激能力
近年来的试验证实,提高泌乳母猪饲粮精氨酸水平或肠道补充精氨酸能够促进哺乳仔猪小肠黏膜生长发育[15-16]。给新生仔猪补充精氨酸可提高其肠道通透性。精氨酸可促进猪小肠上皮细胞增殖[17],在正常条件和应激状态下均能够维护肠道黏膜屏障和保护淋巴细胞,减少热应激造成的肠道黏膜屏障的损伤[18]。最新的研究发现,精氨酸对肠道营养的益处部分归因于精氨酸调控肠道菌群氨基酸代谢及其利用率的结果[19]。断奶仔猪饲粮中补充精氨酸或N-氨基甲酰谷氨酸(合成精氨酸的前体物)在显著提高血浆精氨酸浓度的同时可显著促进小肠的生长和提高黏膜细胞的数量,增加十二指肠、空肠、回肠的绒毛高度以及空肠、回肠的隐窝深度,提高断奶仔猪体重[20]。另外,精氨酸对消化道疾病也有一定的预防和改善作用,例如坏死性肠炎[21]。
饲粮中补充0.83%的精氨酸可显著增强猪机体抗氧化功能[5]。在应激条件下提高精氨酸添加量可以缓解仔猪因应激导致的采食量下降,增强仔猪抗氧化能力,缓解氧化应激对动物生产造成的不利影响[22]。断奶仔猪饲粮中补充精氨酸或N-氨基甲酰谷氨酸可显著提高热休克蛋白70(HSP70)的mRNA表达量和蛋白含量,表明精氨酸有缓解热应激的作用[20]。
2 谷氨酰胺
2.1 谷氨酰胺的代谢途径
谷氨酰胺在动物体内含量丰富,具有多种营养生理功能,因而成为动物功能性氨基酸营养研究的热点,其代谢途径见图4。谷氨酰胺在动物的某些特殊生理状态下,例如应激状态,成为了必需氨基酸,而据研究发现,谷氨酰胺不仅在动物应激状态下发挥重要作用,对正常动物的营养代谢也具有一定的调控作用。谷氨酰胺对氮平衡、肌肉蛋白质代谢、肠黏膜免疫均有一定的益处[23],对营养不良、重大疾病后的恢复也具有明显的改善作用[24]。谷氨酰胺能促进细胞增殖,在机体处于炎症状态下,谷氨酰胺的消耗量高于机体内源产生量。
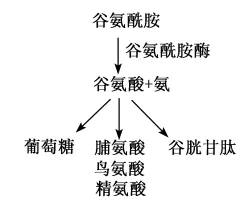
图4 谷氨酰胺的主要代谢途径Fig.4 The major metabolic pathway of glutamine[25]
2.2 谷氨酰胺的营养调控功能
2.2.1 改善动物肠道功能,为营养吸收奠定基础动物肠黏膜细胞能够利用大量的谷氨酰胺作为其能氮来源,以保证正常的肠道功能,促进营养物质吸收。研究发现谷氨酰胺能够显著增加肠道对亮氨酸和脯氨酸的吸收量,显著增加肠道组织游离亮氨酸和脯氨酸量,并显著增加肠道蛋白质的合成量[26]。谷氨酰胺通过调控相关基因表达保护肠黏膜细胞;通过促进还原型谷胱甘肽合成,达到抗氧化作用,维持肠黏膜细胞完整性,减少肠细胞凋亡。饲粮中添加谷氨酰胺可以促进鸡肠道黏膜发育,提高小肠绒毛密度和宽度,增加肠细胞数量[27]。谷氨酰胺可能通过激活mTOR信号通路来促进肠细胞蛋白质合成和肠细胞的生长[28],同时,谷氨酰胺有助于表皮生长因子促进小肠细胞增殖[29],此外,谷氨酰胺的重要中间代谢产物α-酮戊二酸可通过促进mTOR及其下游靶标核糖体S6激酶1(S6K1)和真核细胞翻译起始因子4E结合蛋白1(4E-BP1)的磷酸化来促进小肠上皮细胞蛋白质的合成[30]。紧密连接蛋白是肠细胞紧密连接的主要功能性调节蛋白。缺乏谷氨酰胺时,紧密连接蛋白呈团块状分布于肠上皮细胞胞质内,不能定位在紧密连接处发挥功能;补充谷氨酰胺后,紧密连接蛋白逐渐向细胞膜上转移,定位于膜尖端的紧密连接处,形成完整的紧密连接[31],使肠黏膜形成保护屏障,阻止细菌移位穿过肠细胞,预防肠源性感染,此外,谷氨酰胺也可通过为体内免疫细胞供能来调节肠道免疫。
2.2.2 缓解动物应激,降低生产损失
大量研究表明,谷氨酰胺对生产中的动物应激(热应激、冷应激、运输应激等)具有明显的缓解作用,而且从代谢以及分子水平的研究逐渐揭示了这种营养素能够缓解应激可能的机制。谷氨酰胺可以诱导HSP70表达量增加,而HSP70的高表达可以提高细胞的耐热性,增强动物抵抗热应激的能力。给小白鼠尾静脉注射谷氨酰胺增加了小鼠肝脏、子宫、卵巢组织细胞HSP70的表达,并且HSP70表达量随着谷氨酰胺剂量的加大而增加[32]。谷氨酰胺能够提高动物机体抗氧化相关指标含量,抑制动物在冷应激状态下细胞内产生的有害自由基,因而有助于动物抗冷应激。血清中丙二醛的量常常可以反映机体脂质过氧化的程度,从而间接地反映出细胞受自由基攻击的程度。谷氨酰胺能够降低低温环境下肉羊血清丙二醛水平,从而有效地降低机体细胞受自由基的攻击程度,同时补充谷氨酰胺提高了冷应激肉羊机体血清超氧化物歧化酶(体内的抗氧化剂)活性和总抗氧化能力[33]。外源性补充谷氨酰胺可以防止长时间应激后机体谷氨酰胺含量的下降,具有保护大鼠抵抗运输应激的作用,又可以在某种程度上降低疲劳程度[34]。
2.2.3 调节体内酸碱平衡,维持内环境稳态
动物机体内环境稳态保证了组织、细胞的各种代谢活动的正常进行。在谷氨酰胺酶的作用下,谷氨酰胺被水解生成谷氨酸和氨。当动物机体酸过多时刺激谷氨酰胺酶活性增强,这样,肾脏内肾小管上皮细胞大量的谷氨酰胺水解生成氨结合H+来促进H+从尿中排出,从而降低H+浓度,保证酸碱平衡,防止代谢性酸中毒,维持动物机体内环境稳态,进而利于动物正常生产性能的发挥。
2.2.4 增强动物免疫力
谷氨酰胺可以调节动物体免疫机能,影响动物免疫器官发育。补充谷氨酰胺可以显著提高1周龄肉仔鸡补体C3和免疫球蛋白G(IgG)水平;1~3周龄肉仔鸡胸腺、脾脏指数;血清中免疫球蛋白A(IgA)、免疫球蛋白 M(IgM)和胸腺、法氏囊中白介素-2(IL-2)、白介素-6(IL-6)的水平[35],并且能够显著提高免疫应激仔猪的免疫器官指数[36]。饲粮中添加0.2%、0.4%谷氨酰胺能加快肉仔鸡胸腺和脾脏发育,延缓法氏囊退化;谷氨酰胺可促进肉仔鸡外周血T淋巴细胞的分裂增殖[37]。抗菌肽可在动物免疫中发挥重要作用,而断奶仔猪饲粮中补充谷氨酰胺能够显著促使骨髓、空肠黏膜中抗菌肽PR-39mRNA的表达[38]。
2.2.5 提高动物采食量及产品品质
谷氨酰胺能够调节动物体蛋白质代谢,饲粮中合理补充谷氨酰胺可以提高动物采食量,促进动物生长,提高饲料利用效率,改善动物产品品质。肉仔鸡饲粮中添加0.5%的谷氨酰胺能够显著提高日采食量和日增重[39],添加1.0%、1.5%的谷氨酰胺显著提高了半净膛率,3种添加量的谷氨酰胺均使得皮脂厚和肌间脂肪含量有所降低[40]。用 分 别 含 0(对 照 组)、0.4%、0.7% 和1.0%的饲粮饲喂黄羽肉鸡的试验表明,随着饲粮谷氨酰胺添加水平的提高,56~70d和42~70d的料重比线性降低,1.0%谷氨酰胺组的料重比显著低于对照组[41]。饲粮中补充1.0%的谷氨酰胺能够提高断奶仔猪生长速度和存活率而且能够提高泌乳母猪乳产量[42],并且有提高乳蛋白、乳脂肪、乳糖、非脂固形物含量的趋势[43]。
3 亮氨酸
3.1 亮氨酸的代谢途径
亮氨酸属支链氨基酸中的一种,也是动物必须从饲粮中摄取的必需氨基酸,它在糖代谢、蛋白质的合成和分解,免疫调节等生理生化过程中均发挥重要作用。机体内大部分氨基酸主要在肝脏代谢,但支链氨基酸主要在肌肉中分解代谢,因为肝脏中不存在支链氨基酸转氨酶。由于结构上的相似性,亮氨酸的代谢产物α-酮异己酸和β-羟基-β-甲基丁酸都可调节营养物质代谢(图5)。
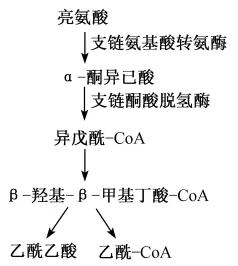
图5 亮氨酸的主要代谢途径Fig.5 The major metabolic pathway of leucine[44]
3.2 亮氨酸的营养调控功能
3.2.1 调节营养物质代谢
亮氨酸通过促进胰岛素和胰高血糖素样肽-1(GLP-1)分泌来降低机体血糖[45]。亮氨酸可依赖于mTOR信号通路或不依赖mTOR信号通路以及刺激某些激素的分泌来促进蛋白质的合成[46-47],可通过调节泛素蛋白酶体途径以及溶酶体介导的自噬途径抑制蛋白质分解[48],但亮氨酸促进蛋白质合成的作用可能具有组织特异性,因为研究发现提高血液中亮氨酸含量可以促进骨骼肌、心肌蛋白质合成[49-50],但不能促进肾脏、脂肪组织蛋白质合成[50],不过也有报道表明在低蛋白基础饲粮中添加亮氨酸可以显著提高断奶仔猪小肠、心脏、肾脏、肝脏、胰脏、脾脏和胃的蛋白质合成速率[51]。以前,关于亮氨酸调控蛋白质合成研究最多的是骨骼肌,除了人类和鼠类上的研究,近年来国内已有在反刍动物上的报道,发现饲粮中每日添加1.0g过瘤胃保护性亮氨酸可以促进绵羊骨骼肌mTOR信号通路中4E-BP1和S6K1的磷酸化[52],进而促进骨骼肌蛋白质合成,此外,亮氨酸代谢产物α-酮异己酸也能显著促进绵羊股二头肌、背最长肌的合成[53]。亮氨酸及其代谢产物可影响绒山羊产绒性能,如不同水平亮氨酸或β-羟基-β-甲基丁酸有提高绒山羊的绒长、绒厚、净绒率的趋势[54]。
3.2.2 调节泌乳功能
进入乳腺的亮氨酸的量总是大于乳中亮氨酸的含量[55-56],这一迹象表明在乳腺中,一部分亮氨酸参与乳蛋白合成;另一部分亮氨酸参与调节了其他物质的合成或发生了氧化代谢。随着奶牛体内可代谢蛋白质的增加,亮氨酸氧化率显著提高[57]。亮氨酸在乳腺内的代谢与其他组织相同,为非必需氨基酸的合成提供碳源和α-氨基氮源[58],具体代谢过程如下:亮氨酸在支链酮酸氨基转移酶(BCAT)的作用下转化为α-酮异己酸,α-酮异己酸在脱氢酶的作用下氧化脱羧生成异戊酰-CoA,异戊酰-CoA进一步转化为β-羟基-β-甲基丁酸-CoA,接着它被2种不同的脱氢酶氧化后,形成乙酰CoA和乙酰乙酸。哺乳动物体内的BCAT分为2种类型:线粒体型(mBCAT)和胞质型(cBCAT)[59],De Santiago等[60]在小鼠乳腺中发现了mBCAT,而且在泌乳期,大鼠乳腺内mBCAT的基因表达、蛋白质合成及其活性至少要非泌乳期高10倍。此外,研究表明只有处于泌乳期的大鼠乳腺上皮细胞才表达mBCAT基因,而且mBCAT的活性与产奶量相关[61]。体外培养基中去除亮氨酸可显著降低奶牛乳腺上皮细胞β-乳球蛋白的合成,以及mTOR下游靶标4E-BP1和S6K1的磷酸化水平,表明亮氨酸具有影响乳蛋白合成与调节乳蛋白合成信号通路的双重作用[62],也有报道表明提高亮氨酸循环水平有利于泌乳[63]。
3.2.3 其他功能
亮氨酸具有调节激素分泌、酶活性、免疫的作用,例如,它可以通过促进瘦素的合成与分泌来调节动物的饱腹感[64]。亮氨酸及其代谢产物β-羟基-β-甲基丁酸可通过提高血清中IgG、IgM的含量,降低血清中可溶性CD4和CD8的含量来增强机体特异性免疫机能[54]。亮氨酸可独立于胰岛素直接长期调控胰腺α-淀粉酶的分泌[65]。
4 色氨酸
4.1 色氨酸的代谢途径
色氨酸属于芳香族氨基酸,是一种兼性生糖生酮氨基酸,也是单胃动物和幼龄反刍动物的必需氨基酸。越来越多的研究证实了色氨酸及其代谢产物对动物的采食量、繁殖、免疫、神经功能、抗氧化、营养代谢等方面均产生调节作用[66]。色氨酸是5-羟色胺、烟酰胺腺嘌呤二核苷酸(NAD)、烟酸、褪黑激素等重要物质的前体物,其中5-羟色胺通过与其受体结合调节动物中枢神经系统;NAD是一种重要的氧化还原辅酶,在呼吸链中发挥重要作用;烟酸具有调节消化道健康和血液循环的作用;褪黑激素具有调节内分泌、抗氧化等多种功能(图6)。
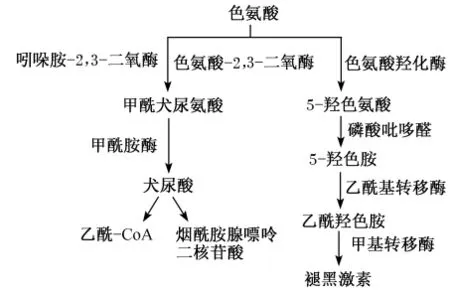
图6 色氨酸的主要代谢途径Fig.6 The major metabolic pathway of tryptophan[66]
4.2 色氨酸的营养调控功能
4.2.1 调控采食和营养物质代谢
人们首先发现色氨酸具有调节动物采食量的作用。饲粮中添加适当水平的色氨酸可调节动物采食量[67],这种作用可能与色氨酸及其代谢产物5-羟色胺参与神经调节以及刺激调控食欲的相关激素的合成与分泌有关。例如,Zhang等[68]发现色氨酸可通过促进胃肠激素(ghrelin)的分泌和胃肠黏膜ghrelin mRNA的表达来增进断奶仔猪食欲和促进采食。色氨酸参与调节体内蛋白质代谢,试验表明,在蛋鸡饲粮中添加适宜水平的色氨酸能够促进机体的蛋白质代谢,提高产蛋性能[69]。饲粮色氨酸对生长猪的氮沉积也具有显著影响[70]。魏宗友等[71]研究发现,5~10周龄扬州鹅饲粮中添加适宜水平的色氨酸可显著提高胸肌率。色氨酸调节蛋白质合成与降解可能与其参与调节蛋白质合成与降解的信号途径有关,在鹅上的研究表明,色氨酸是通过上调参与调控蛋白质合成基因的mRNA表达量、蛋白合成量及磷酸化水平,降低促进蛋白质降解基因表达来促进腿肌蛋白质沉积的[72],此外,它可能还可通过调节体内激素分泌,例如提高血液中胰岛素样生长因子-Ⅰ(IGF-Ⅰ)水平来调节蛋白质代谢[73]。除了蛋白质代谢,色氨酸也参与调节体内脂肪代谢,试验观察到在显著降低蛋鸡肝脂率和腹脂率的同时能显著提高血清极低密度脂蛋白胆固醇含量和腹脂组织中激素敏感脂肪酶活性[74]。
4.2.2 调节神经和免疫功能
色氨酸通过其代谢产物5-羟色胺的中枢神经调节作用来减少动物因应激发生的争斗行为[75]。饲粮中添加0.04%的色氨酸可显著改善蛋雏鸭提高机体的抗氧化能力,促进免疫器官的发育,提高其生长性能[76]。肉鸡饲粮中补充色氨酸和精氨酸能显著提高血α-干扰素、γ-干扰素以及免疫球蛋白含量,增强肉鸡抵抗传染性法氏囊病的能力[77]。色氨酸缺乏可以引起扬州鹅脾脏组织淋巴细胞生长受抑制并发生凋亡,适当补充色氨酸可促进脾脏组织淋巴细胞的增殖与分化,从而有利于改善扬州鹅免疫力[72]。近年来还发现色氨酸可不依赖于维生素B6独立调节断奶仔猪的体内的免疫反应[78],提高仔猪免疫力。色氨酸的一些代谢产物也参与免疫调节,例如羟基喹啉可通过补充白细胞内的NAD来缓解氧化应激造成的NAD的减少来调节免疫[79]。
4.2.3 色氨酸代谢产物对反刍动物的作用
色氨酸的代谢产物褪黑激素可通过调节其他激素分泌及皮肤脱碘酶活性来促进绒山羊产绒[80-81],这种作用还可能与褪黑激素调节参与调节调控产绒的基因表达和信号转导有关,例如,褪黑激素可提高参与毛囊生长的信号转导的Wnt10b基因在绒山羊皮肤组织中毛囊休止期和退行期的表达量[82],通过调节PDGFA基因表达缩短毛囊生长周期[83]。除了促进产绒,褪黑激素对免疫和繁殖也有调控作用[84-85]。色氨酸的代谢产物烟酸在反刍动物上研究较多,烟酸在调节瘤胃微生物菌群和消化率、酮症的预防、抗应激以及产奶量和乳成分含量与产量等方面均可发挥积极作用[86]。
5 苏氨酸
5.1 苏氨酸的代谢
苏氨酸是动物体内的重要的必需氨基酸之一,体内苏氨酸的在不同的酶的作用下有不同的代谢途径(图8),但主要是在苏氨酸脱氢酶作用下生成丙酮酸、甘氨酸和乙酰辅酶A,在猪上的研究表明,苏氨酸的这种分解代谢仅在肝脏和胰腺中发生[87]。在饥饿条件下,苏氨酸可在苏氨酸脱水酶的作用下生成α-酮丁酸和氨[88]。除了苏氨酸脱氢酶和脱水酶之外,苏氨酸可受到苏氨酸醛缩酶的作用。
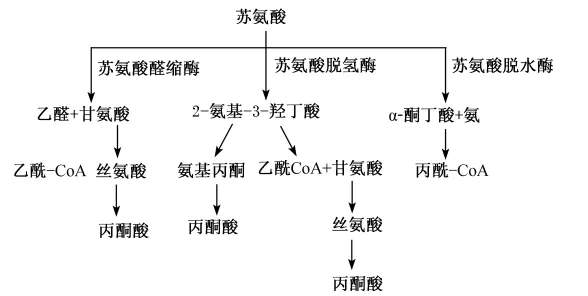
图7 苏氨酸的主要代谢途径Fig.7 The major metabolic pathway of threonine[87]
5.2 苏氨酸的营养调控功能
5.2.1 改善低蛋白质饲粮下的动物生产性能
饲粮中添加适宜量的苏氨酸可显著提高动物采食量和生产性能,尤其是在低蛋白质饲粮条件下补充苏氨酸对动物生产性能具有更明显的改善作用。饲粮补充添加0.1%~0.4%的苏氨酸能极显著提高肉兔平均日采食量[89],在肉仔鸡饲粮中添加0.04%的苏氨酸能明显提高生产性能和肉鸡的均匀度。在低蛋白质饲粮条件下,添加较高水平的苏氨酸能改善肉鸡胴体组成,提高腹脂、胸脂含量以及胴体与屠体重的比值[90];添加0.1%~0.3%的苏氨酸可显著提高蛋鸡饲料利用率、产蛋率和蛋壳质量,促进鸡蛋蛋白质的沉积[91];补充苏氨酸显著增加鸭羽毛质量和长度[92]。生产实际中必须考虑苏氨酸的适宜添加量,因为过量的苏氨酸很可能导致动物采食量和生产性能的下降[93],其机理到目前还未阐明。
5.2.2 免疫调节作用
当动物处于免疫应激条件下,免疫球蛋白的大量合成易引起饲粮苏氨酸的缺乏和体蛋白质的动员[94],因此饲粮中补充苏氨酸有利于改善动物的免疫性能,提高抗病能力。据报道,饲粮中补充苏氨酸可显著提高小鼠体内循环抗体的含量和脾T淋巴细胞的转化功能[95]。补充苏氨酸对感染伪狂犬病毒猪空肠上皮细胞先天性免疫功能具有分子表达水平的调控作用[96]。
5.2.3 调节蛋白质合成
饲粮苏氨酸可通过调节小肠黏蛋白的合成来影响断奶仔猪肠道的健康和肠道功能[97]。苏氨酸是小肠合成黏蛋白所必需的营养素[98],而黏蛋白具有维持小肠屏障的功能,当小肠供给的苏氨酸不足时导致肠黏膜屏障遭到损伤最终可能会导致幼龄动物坏死性肠炎的发生[99]。除了小肠黏蛋白,饲粮添加不同水平苏氨酸对断奶仔猪肝脏中蛋白质水平也有显著影响[100]。
6 小 结
综上所述,氨基酸作为合成体内蛋白质前体物,同时又因某些氨基酸是一些功能性物质的前体物而具有了生物活性功能。精氨酸、谷氨酰胺、亮氨酸、色氨酸、苏氨酸在整体水平可调控动物的采食量、生产性能、饲料转化率、健康状况等;在组织水平可调节组织生长发育和功能;在代谢水平可调节酶活性、蛋白质等营养物质的代谢、免疫物质的合成及神经内分泌;在分子水平参与调控基因的表达、蛋白质的合成及信号通路。这些功能使得具有生物活性的氨基酸逐渐成为了研究热点,未来的研究有望系统地解析这些氨基酸的具体作用机理,同时,注意一些氨基酸(如精氨酸、谷氨酰胺、脯氨酸等)家族中不同氨基酸之间的协调作用机制研究。确定其在各种动物不同生长阶段或不同生理状态下发挥某种调节作用的适宜添加量,开发具有动物生产改良与动物保健的活性氨基酸及其类似物的添加剂,从而提高养殖业的生产效率。
[1] WU G,BAZER F W,DAVIS T A,et al.Arginine metabolism and nutrition in growth,health and disease[J].Amino Acids,2009,37(1):153-168.
[2] JOBGEN W S,FRIED S K,FU W J,et al.Regulatory role for the arginine-nitric oxide pathway in metabolism of energy substrates[J].The Journal of Nutritional Biochemistry,2006,17(9):571-588.
[3] WU G,MORRIS S M,Jr.Arginine metabolism:nitric oxide and beyond[J].Biochemical Journal,1998,336(1):1-17.
[4] 杨静,谢明,侯水生,等.精氨酸调控畜禽采食量的机制及其影响因素[J].动物营养学报,2012,24(4):612-616.
[5] 吴琛,刘俊锋,孔祥峰,等.饲粮精氨酸与丙氨酸对环江香猪肉质、氨基酸组成及抗氧化功能的影响[J].动物营养学报,2012,24(3):528-533.
[6] 方勇军.精氨酸对肉鸭生长性能、免疫机能、胴体品质和血液脂质的影响[D].硕士学位论文.武汉:武汉工业学院,2009.
[7] MC KNIGHT J R,SATTERFIELD M C,JOBGEN W S,et al.Beneficial effects of L-arginine on reducing obesity:potential mechanisms and important implications for human health[J].Amino Acids,2010,39(2):349-357.
[8] KIM S W,WU G.Regulatory role for amino acids in mammary gland growth and milk synthesis[J].Amino Acids,2009,37(1):89-95.
[9] MATEO R D,WU G,MOON H K,et al.Effects of dietary arginine supplementation during gestation and lactation on the performance of lactating primiparous sows and nursing piglets[J].Journal of Animal Science,2008,86(4):827-835.
[10] VICINI J L,CLARK J H,HURLEY W L,et al.Effects of abomasal or intravenous administration of arginine on milk production,milk composition,and concentrations of somatotropin and insulin in plasma of dairy cows[J].Journal of Dairy Science,1988,71(3):658-665.
[11] CHEW B P,EISENMAN J R,TANAKA T S.Arginine infusion stimulates prolactin,growth hormone,insulin,and subsequent lactation in pregnant dairy cows[J].Journal of Dairy Science,1984,67(11):2507-2518.
[12] DOEPEL L,LAPIERRE H.Deletion of arginine from an abomasal infusion of amino acids does not decrease milk protein yield in Holstein cows[J].Journal of Dairy Science,2011,94(2):864-873.
[13] 徐柏林,王梦芝,张兴夫,等.精氨酸水平对奶牛乳腺上皮细胞体外生长及κ-酪蛋白基因表达的影响[J].动物营养学报,2012,24(5):851-857.
[14] 徐柏林.精氨酸对乳腺上皮细胞中酪蛋白合成的影响及其调控机制[D].硕士学位论文.扬州:扬州大学,2012.
[15] PUIMAN P J,STOLL B,VAN GOUDOEVER J B,et al.Enteral arginine does not increase superior mesenteric arterial blood flow but induces mucosal growth in neonatal pigs[J].The Journal of Nutrition,2011,141(1):63-70.
[16] 郭长义,蒋宗勇,李职,等.泌乳母猪饲粮精氨酸水平对哺乳仔猪小肠黏膜发育的影响[J].动物营养学报,2010(4):870-878.
[17] 聂新志,蒋宗勇,林映才,等.精氨酸和谷氨酰胺对猪小肠上皮细胞增殖的影响及机理探讨[J].中国农学通报,2012(2):1-5.
[18] 张灿菲.精氨酸对急性热应激鸡肠道黏膜免疫的影响[D].硕士学位论文.武汉:华中农业大学,2008.
[19] DAI Z L,LI X L,XI P B,et al.Regulatory role for L-arginine in the utilization of amino acids by pig smallintestinal bacteria[J].Amino Acids,2012,43(1):233-244.
[20] WU X,RUAN Z,GAO Y,et al.Dietary supplementation with L-arginine or N-carbamylglutamate enhances intestinal growth and heat shock protein-70expression in weanling pigs fed a corn-and soybean meal-based diet[J].Amino Acids,2010,39(3):831-839.
[21] KUL M,VURUCU S,DEMIRKAYA E,et al.Enteral glutamine and/or arginine supplementation have favorable effects on oxidative stress parameters in neonatal rat intestine[J].Journal of Pediatric Gastroenterology and Nutrition,2009,49(1):85-89.
[22] 郑萍.氧化应激对仔猪精氨酸代谢和需求特点的影响及机制研究[D].博士学位论文.雅安:四川农业大学,2010.
[23] VAN ACKER B A,VON MEYENFELDT M F,SOETERS P B.Glutamine as a key ingredient in protein metabolism[J].Ned Tijdschr Geneeskd,1999,143(38):1904-1908.
[24] VAN DERHULST R R,VON MEYENFELDT M F,SOETERS P B.Glutamine:an essential amino acid for the gut[J].Nutrition,1996,12(11/12):78-81.
[25] WATFORD M.Glutamine metabolism and function in relation to proline synthesis and the safety of glutamine and proline supplementation[J].The Journal of Nutrition,2008,138(10):2003-2007.
[26] 叶元土,王永玲,蔡春芳,等.谷氨酰胺对草鱼肠道L-亮氨酸、L-脯氨酸吸收及肠道蛋白质合成的影响[J].动物营养学报,2007,20(1):28-32.
[27] MURAKAMI A E,SAKAMOTO M I,NATALI M R,et al.Supplementation of glutamine and vitamin E on the morphometry of the intestinal mucosa in broiler chickens[J].Poultry Science,2 0 0 7,8 6(3):4 8 8-4 9 5.
[28] 席鹏彬,蒋宗勇,戴兆来,等.L-谷氨酰胺对猪肠上皮细胞蛋白质周转的调控[C]//第六次全国饲料营养学术研讨会论文集.杨凌:中国畜牧兽医学会,2010.
[29] KO T C,BEAUCHAMP R D,TOWNSEND C M,Jr,et al.Glutamine is essential for epidermal growth factor-stimulated intestinal cell proliferation[J].Surgery,1993,114(2):147-153.
[30] YAO K,YIN Y,LI X,et al.Alpha-ketoglutarate inhibits glutamine degradation and enhances protein synthesis in intestinal porcine epithelial cells[J].Amino Acids,2012,42(6):2491-2500.
[31] 崔巍,闻颖,董亚珞,等.谷氨酰胺对体外培养肠上皮细胞屏障通透性的影响[J].世界华人消化杂志,2008,33:3729-3733.
[32] 张志宏,姜忠玲,曹荣峰,等.谷氨酰胺诱导热休克蛋白70表达的研究[J].中国奶牛,2 0 0 9(4):7-1 0.
[33] 高福久.谷氨酰胺和L-肉碱对肉羊抗低温应激能力影响的研究[D].硕士学位论文.大庆:黑龙江八一农垦大学,2010.
[34] 吴艳.大鼠的运输应激及谷氨酰胺的抗应激研究[D].硕士学位论文.南京:南京农业大学,2008.
[35] 张敏,邹晓庭,孙亚丽,等.谷氨酰胺对1~3周龄肉仔鸡免疫功能的影响[J].畜牧兽医学报,2009(10):1494-1498.
[36] 陈静,刘显军,张飞,等.谷氨酰胺对免疫应激仔猪免疫器官指数的影响[J].中国兽医杂志,2010(9):3-5.
[37] 周联高,章世元,刘艳芬,等.谷氨酰胺对肉仔鸡生产性能及免疫机能的影响[J].动物营养学报,2008(3):305-310.
[38] 赵玉蓉,王红权,贺建华,等.谷氨酰胺对断奶仔猪抗菌肽PR-39mRNA的表达调控[J].动物营养学报,2009(4):567-572.
[39] 董金格,万晓媛,邹晓庭,等.外源添加谷氨酰胺对肉仔鸡生长性能及抗氧化指标的影响[J].饲料工业,2008(22):4-6.
[40] 董金格,胡家澄,邹晓庭,等.外源添加谷氨酰胺对肉仔鸡生长性能和胴体组成的影响[J]中国饲料,2009(1):33-35.
[41] 黄冠庆,林红英,黄晓亮,等.谷氨酰胺对黄羽肉鸡生长、抗氧化力及肉品质的影响[J].中国畜牧杂志,2010(21):60-64.
[42] WU G,BAZER F W,JOHNSON G A,et al.Triennial growth symposium:important roles for L-glutamine in swine nutrition and production[J].Journal of Animal Science,2011,89(7):2017-2030.
[43] 秦江帆,蒋宗勇,林映才,等.谷氨酰胺对母猪泌乳性能、氨基酸代谢的影响[C]//中国畜牧兽医学会动物营养学分会第十次学术研讨会论文集.杭州:中国农业科学技术出版社,2008.
[44] VIANNA D,TEODORO G F R,TORRES-LEAL F L,et al.Protein synthesis regulation by leucine[J].Brazilian Journal of Pharmaceutical Sciences,2010,46(1):29-36.
[45] 王觐,薛长勇,徐庆,等.亮氨酸对血糖的影响及其相关机制研究[J].军医进修学院学报,2012(2):132-134.
[46] ANTHONY J C,LANG C H,CROZIER S J,et al.Contribution of insulin to the translational control of protein synthesis in skeletal muscle by leucine[J].A-merican Journal of Physiology:Endocrinology and Metabolism,2002,282(5):1092-1101.
[47] 毛湘冰,黄志清,陈小玲,等.亮氨酸调节哺乳动物骨骼肌蛋白质合成的研究进展[J].动物营养学报,2011(5):709-714.
[48] 毛湘冰,余冰,陈代文,等.亮氨酸调节动物骨骼肌蛋白质分解的研究进展[J].中国畜牧杂志,2011(17):73-76.
[49] ESCOBAR J,FRANK J,SURYAWAN A,et al.Cardiac and skeletal muscle protein synthesis and activation of translation initiation factors are stimulated by leucine,but not isoleucine or valine,in neonatal pigs[J].Journal of dairy science,2005,88:239-239.
[50] ESCOBAR J,NGUYEN H V,DAVIS T A.Differential effects of leucine on translation initiation factor activation and protein synthesis in skeletal muscle,renal and adipose tissues of neonatal pigs[J].Journal of Dairy Science,2007,90:441-441.
[51] 王彬,李奇,亮氨酸对断奶仔猪日增重和组织器官蛋白质合成速率的影响[J].江苏农业科学,2012(5):166-167.
[52] 桑丹,孙海洲,郭俊清,等.过瘤胃保护性亮氨酸对绵羊骨骼肌哺乳动物雷帕霉素靶蛋白(mTOR)信号传导通路关键因子的影响[J].动物营养学报,2011(1):61-65.
[53] 桑丹.亮氨酸及α-酮异己酸钙对绵羊机体蛋白质合成的影响研究[D].硕士学位论文.呼和浩特:内蒙古农业大学,2009.
[54] 郭俊清.亮氨酸及β-羟基-β-甲基丁酸钙对绒山羊免疫机能和生产性能影响的研究[D].硕士学位论文.呼和浩特:内蒙古农业大学,2009.
[55] BEQUETTE B J,BACKWELL F R.Amino acid supply and metabolism by the ruminant mammary gland[J].Proceedings of the Nutrition Society,1997,56(2):593-605.
[56] MEPHAM T B.Amino acid utilization by lactating mammary gland[J].Journal of Dairy Science,1982,65(2):287-298.
[57] BEQUETTE B J,METCALF J A,WRAY-CAHEN D,et al.Leucine and protein metabolism in the lactating dairy cow mammary gland:responses to supplemental dietary crude protein intake[J].Journal of Dairy Research,1996,63(2):209-222.
[58] WOHLT J E,CLARK J H,DERRIG R G,et al.Valine,leucine,and isoleucine metabolism by lactating bovine mammary tissue[J].Journal of Dairy Science,1977,60(12):1875-1882.
[59] HALL T R,WALLIN R.,REINHART G D,et al.Branched chain aminotransferase isoenzymes.Purification and characterization of the rat brain isoenzyme[J].The Journal of Biological Chemistry,1993,268(5):3092-3098.
[60] DE SANTIAGO S,TORRES N,SURYAWAN A,et al.Regulation of branched-chain amino acid metabolism in the lactating rat[J].The Journal of Nutrition,1998,128(7):1165-1171.
[61] TOVAR A R,BECERRIL E,HERNANDEZ-PANDO R,et al.Localization and expression of BCAT during pregnancy and lactation in the rat mammary gland[J].American Journal of Physiology:Endocrinology and Metabolism,2001,280(3):480-488.
[62] MOSHEL Y,RHOADS R E,BARASH I.Role of amino acids in translational mechanisms governing milk protein synthesis in murine and ruminant mammary epithelial cells[J].Journal of Cellular Biochemistry,2006,98(3):685-700.
[63] LEI J,FENG D,ZHANG Y,et al.Regulation of leu-cine catabolism by metabolic fuels in mammary epithelial cells[J].Amino Acids,2 0 1 2,4 3(5):2 1 7 9-2 1 8 9.
[64] LI F,YIN Y,TAN B,et al.Leucine nutrition in animals and humans:mTOR signaling and beyond[J].A-mino Acids,2011,41(5):1185-1193.
[65] 于红霞,于志鹏,刘凯,等.十二指肠灌注亮氨酸对山羊胰腺外分泌功能的影响[J].动物营养学报,2011(9):1513-1518.
[66] YAO K,FANG J,YIN Y L,et al.Tryptophan metabolism in animals:important roles in nutrition and health[J].Frontiers in Bioscience(Scholar edition),2011,3:286-297.
[67] 马玉娥,占秀安,朱巧明,等.饲粮色氨酸水平对黄羽肉种鸡生产性能、抗氧化功能及血清生化指标的影响[J].动物营养学报,2011(12):2177-2182.
[68] ZHANG H,YIN J,LI D,et al.Tryptophan enhances ghrelin expression and secretion associated with increased food intake and weight gain in weanling pigs[J].Domestic Animal Endocrinology,2 0 0 7,3 3(1):4 7-6 1.
[69] 饶巍,王玥,周斌,等.日粮色氨酸水平对蛋鸡生产性能及蛋白质代谢的影响[J].中国畜牧杂志,2011(15):38-41.
[70] 任建波,赵广永,李元晓,等.日粮色氨酸水平对生长猪的氮利用效率、血浆类胰岛素生长因子-Ⅰ、生长激素及胰岛素的影响[J].动物营养学报,2007(3):264-268.
[71] 魏宗友,王洪荣.色氨酸对5~10周龄扬州鹅生长性能和屠宰性能的影响[J].中国饲料,2012(12):29-34.
[72] 魏宗友.色氨酸对鹅组织蛋白质代谢调控机制和免疫功能的影响[D].博士学位论文.扬州:扬州大学,2012.
[73] 张华伟,尹靖东,周玄,限饲和自由采食条件下色氨酸对断奶仔猪血清IGF-1浓度的影响[J].中国畜牧杂志,2006(23):19-22.
[74] 周斌,李慧,邹晓庭,等.色氨酸对产蛋鸡脂肪代谢的影响[J].中国畜牧杂志,2011(7):50-53.
[75] 刘化伟,石宝明.色氨酸对仔猪应激行为的影响[J].饲料博览,2010(11):14-16.
[76] 刘肖挺,王安,杨小然,等.色氨酸对蛋雏鸭生长性能、抗氧化功能及免疫器官发育的影响[J].饲料工业,2012(10):5-8.
[77] EMADI M,JAHANSHIRI F,KAVEH K,et al.Nutrition and immunity:the effects of the combination of arginine and tryptophan on growth performance,serum parameters and immune response in broiler chickens challenged with infectious bursal disease vaccine[J].Avian Pathology,2011,40(1):63-72.
[78] MATTE J J,LEFLOC’H N,PRIMOT Y,et al.Interaction between dietary tryptophan and pyridoxine on tryptophan metabolism,immune responses and growth performance in post-weaning pigs[J].Animal Feed Science and Technology,2011,170(3):256-264.
[79] MOFFETT J R,NAMBOODIRI M A.Tryptophan and the immune response[J].Immunology & Cell Biology,2003,81(4):247-265.
[80] 程建波,张子军,章孝荣,等.褪黑激素对绒山羊绒毛生长的影响及其作用机理研究进展[J].贵州农业科学,2011(3):166-168,171.
[81] 岳春旺,孙茂红,朱晓萍,等.外源褪黑激素对绒山羊皮肤脱碘酶活性的影响[J].西北农林科技大学学报:自然科学版,2011(6):49-54.
[82] 丽春,张文广,常子丽,等.Wnt10b参与外源褪黑激素促进绒山羊绒毛生长的研究[J].中国畜牧杂志,2012(7):5-8.
[83] 丽春,张文广,杨文柱,等.褪黑激素对PDGFA基因在绒山羊皮肤毛囊中表达模式的影响[J].中国农业科学,2012(10):2031-2039.
[84] 赵国先,李娜,宋海彬,等.褪黑素对动物的免疫调节作用[J].中国饲料,2008(15):11-14.
[85] 郭玉强,蔡国宝,刘艳婷,等.褪黑激素对绵羊季节性繁殖的调控机理[J].甘肃畜牧兽医,2008(1):43-46.
[86] 蒋亚军,周凌云,赵芸君,等.烟酸在反刍动物营养中的研究进展[J].中国畜牧兽医,2010(7):9-14.
[87] LEFLOC’H N,THIBAULT J N,SEVE B.Tissue localization of threonine oxidation in pigs[J].British Journal of Nutrition,1997,77(4):593-603.
[88] KlDD M T,KERR B J.L-threonine for poultry:a review[J].The Journal of Applied Poultry Research,1996(5):358-367.
[89] 张艳蕾,王雪鹏,王春阳,等.饲粮苏氨酸水平对断奶至2月龄肉兔生长性能、免疫功能和血清生化指标的影响[J].动物营养学报,2011(4):703-708.
[90] 龚利敏,李德发,陈刚.苏氨酸对肉鸡生产性能和产肉量的影响[C]//第四届全国饲料营养学术研讨会论文集.南京:中国农业科学技术出版社,2002.
[91] 刘国花,邹晓庭,谢正军,等.低蛋白饲粮添加苏氨酸对蛋鸡生产性能及蛋品质的影响[J].中国家禽,2012(9):33-36.
[92] 郭锋,张丽,侯水生,等.不同粗蛋白和苏氨酸水平对生长前期北京鸭羽毛发育的影响[J].畜牧兽医学报,2012(1):65-70.
[93] EDMONDS M S,GONYOU H W,BAKER D H.Effect of excess levels of methionine,tryptophan,arginine,lysine or threonine on growth and dietary choice in the pig[J].Journal of Animal Science,1987,65(1):179-185.
[94] 林燕.日粮苏氨酸水平对感染伪狂犬病毒雄鼠免疫功能和繁殖性能的影响[D].硕士学位论文.雅安:四川农业大学,2009.
[95] 刘华忠,游金明,钟杰平.苏氨酸对小鼠免疫功能的影响[J].河南科技大学学报:自然科学版,2006(5):72-74,109-110.
[96] 韩国全,余冰,陈代文,等.苏氨酸对体外培养感染伪狂犬病毒猪空肠上皮细胞免疫相关基因表达的影响[J].动物营养学报,2012(3):487-496.
[97] 王旭,谯仕彦,印遇龙,等.苏氨酸对断奶仔猪小肠黏膜和黏液蛋白质合成的影响[C]//中国畜牧兽医学会2006学术年会论文集(上册).北京:中国畜牧兽医学会,2006.
[98] LAW G K,BERTOLO R F,ADJIRI-AWERE A,et al.Adequate oral threonine is critical for mucin production and gut function in neonatal piglets[J].American Journal of Physiology:Gastrointestinal and Liver Physiology,2007,292(5):1293-1301.
[99] PUIMAN P J,JENSEN M,STOLL B,et al.Intestinal threonine utilization for protein and mucin synthesis is decreased in formula-fed preterm pigs[J].The Journal of Nutrition,2011,141(7):1306-1311.
[100]侯永清,吕民主,呙于明,等.早期断奶仔猪日粮中蛋氨酸、苏氨酸水平对机体蛋白质代谢的影响[J].饲料研究,2001(7):7-8.
猜你喜欢
动物营养学报(2022年12期)2023-01-05
北方牧业(2022年9期)2022-11-22
微生物学杂志(2022年5期)2022-03-09
家畜生态学报(2020年7期)2020-07-14
中国饲料(2019年5期)2019-01-10
分析化学(2018年4期)2018-11-02
猪业科学(2017年2期)2017-03-30
广东饲料(2016年1期)2016-12-01
湖南饲料(2015年1期)2015-04-07
中国饲料(2015年12期)2015-01-25
