
高寒沙区6种生态恢复树种蒸腾实测1)
2013-08-08 07:22王学全刘君梅尹书乐杨恒华
东北林业大学学报 2013年9期
陈 琦 王学全 刘君梅 尹书乐 杨恒华
(中国林业科学研究院荒漠化研究所,北京,100091) (青海省沙珠玉治沙站)
近几十年来,在自然环境变化和人为干扰的双重作用下,青海共和盆地的荒漠化程度进一步加深。随着全球变暖,共和盆地近几十年来出现了气温逐步升高、降水量微弱增加、蒸发量增大的气候变化趋势,共和盆地气候将向干热方向演化,干旱程度将会加重[1-3]。在人工植被恢复建植过程中,需要考虑水分承载力,以使恢复后的植被群落向健康稳定的方向发展。
植物种的选择是植被恢复和重建的第一步,也是人工植物群落结构调控的手段[4]。在干旱、半干旱地区,水资源短缺问题日益严重,水分成为造林的主要限制因子[5-6]。面对水资源不足的现状,选择低耗水树种进行人工植被恢复是林学家、生态学家共同关注的问题[7-9]。蒸腾是植物耗水特征的主要方式,在土壤—植物—大气连续体(SPAC)水热传输过程中占有极为重要的地位[10]。在以往对共和盆地植被恢复树种的研究中,多侧重于植物叶性因子的差异[11]、适宜树种造林技术[12]以及生态恢复对物种多样性的影响[13]等方面的探讨,而对植物蒸腾耗水缺乏定量研究。因此,选用适宜方法及时开展这项工作具有理论和现实上的必要性。测定单株植物蒸腾耗水的方法多种多样。目前,国外学者多采用液流法,该法可分析植物蒸腾与自身生理因子的关系,但当树干液流较小时,该方法不准确[14];国内学者则主要采用光合系统测定仪法,该法可探讨植物蒸腾速率与微气象因子间的响应,但所测数据不能用于精确推算实际耗水量[15]。笔者采用快速称质量法,该方法简单易行,定量较准确,适宜于不同树种、不同时间之间的比较研究,且离体叶片处于自然环境中,能较好地反映环境对蒸腾的影响[16]。笔者通过研究分析了高寒沙区6种生态恢复树种的蒸腾速率日动态、瞬时蒸腾量日动态及日蒸腾量,并对不同方位植物叶片的蒸腾差异进行了对比,以期为高寒沙区树种选择和植被恢复提供指导和依据。
1 研究地概况
研究区位于青海共和盆地高寒沙区沙珠玉治理示范区,地理坐标 98°46'~101°22'E,35°27'~ 36°56'N,海拔2 871~3 000 m,属高寒干旱荒漠和半干旱草原过渡区域。年均气温2.4℃,无霜期91 d,年日照时间 2 772.2 h,总辐射量 662.87 kJ·cm-2,生理辐射311.54 kJ·cm-2。年降水量250 ~420 mm,季节分配不均,5—9月份降水量占全年的89.2%,年蒸发量1400~2400 mm。研究区原来的流动沙丘,通过设置机械沙障后直播油蒿、柠条、丘间地乌柳造林,建立起固沙植被,经过长期演变,植被由最初的单一人工种植灌木、半灌木演变到出现1年生和多年生草本,土壤开始出现结皮。土壤以风沙土为主,分为流动风沙土、半固定风沙土及固定风沙土[17-18]。
2 材料与方法
供测植物种选取治沙示范区主要植被恢复物种,包括垂榆(Ulmus pumila var.pendula)、小叶杨(Populus simonii)、柠条(Caragana korshinskii)、沙棘(Hippophae rhamnoides)、乌柳(Salix cheilophila)、沙柳(Salix psammophila),其中柠条、沙棘生长于固定沙丘,其余树种生长于丘间地,供测植株株龄均为10 a左右,其中小叶杨为20 a,各植物种均处于其生长壮年期,测定各植株蒸腾速率并计算日蒸腾量。
蒸腾速率的测定采用快速称质量法,时间为七八月份,选择典型晴天,07:00—19:00每间隔2 h测定1次,共7个时间点,每个树种共测定4次。测定时,在野外选择无病虫害正常生长的植物标准样株,于冠层中部东、北、西、南4个方向各摘取一定数量的叶片(其中小叶杨2片,垂榆3片,沙棘和沙柳各取10片,柠条取3片复叶,乌柳取3个小枝),置于千分计精确天平的田间防风罩内进行快速称质量,完成后,仍将叶片置于野外自然环境中,间隔3 min再次称质量。由于叶片均自叶柄基部摘取,创伤面小,对叶片失水影响较小,故计算2次称质量差值可为该时段内叶片的蒸腾失水量。将所测叶片带回室内进行扫描,利用Image-pro plus 6.0软件计算单叶面积。蒸腾速率计算公式为:
Tr=ΔW/ΔT/Sp。
式中:Tr为蒸腾速率(μg·cm-2·s-1);ΔW 为 2 次称质量差值(μg);ΔT为2次称质量间隔时间(s);Sp为单次称质量叶片总面积(cm2)。
在野外计算实测植物的总叶数量,与单叶面积相乘得到总叶面积。取各时刻蒸腾速率的平均值为植株白天蒸腾速率的平均值。日蒸腾量计算公式为:
Ew=Tm×T×St。
式中:Ew为日蒸腾量(μg);Tm为日平均蒸腾速率(μg·cm-2·s-1);T 为全天测定时长(s),即12 h;St为单株植物总叶面积(cm2)。
3 结果与分析
3.1 植物蒸腾速率日动态
供试树种的蒸腾速率日变化如图1所示,所有树种蒸腾速率变化均呈先升后降的趋势,其中垂榆、小叶杨、沙棘、沙柳日进程曲线为单峰型,没有出现明显的蒸腾午休现象,可归为非蒸腾午休型[19],柠条、乌柳日进程曲线呈双峰型,且第二个峰值较第一个峰值高,但双峰曲线具有不典型性,峰谷不明显。各树种蒸腾速率峰值出现时间不同,其中垂榆和沙棘均在11:00达到峰值,如表1所示,其峰值分别为为 5.17、8.71 μg·cm-2·s-1,随后蒸腾速率开始下降,且相比其上升趋势,下降趋势更为平缓,是植物为了保持自身水分而采取的一种保护措施,这与段爱国等[20]对苏门答腊金合欢、木豆的研究相一致。沙柳于正午13:00达到其蒸腾速率峰值10.35 μg·cm-2·s-1,从全天来看,09:00—17:00,其蒸腾速率均处在较高值,这与该时段光照充足、气温较高、植物自身气孔扩散阻力较低相关。小叶杨的峰值出现时间较晚,于 15:00 达到 6.18 μg·cm-2·s-1,表现出明显的蒸腾作用滞后性,可能与其自身生理结构对干旱环境的适应相关。乌柳的蒸腾速率在清晨上升较快,于09:00达到第一个峰值,为全天次高值,随后出现微弱下降,呈现不明显的蒸腾午休现象,在正午13:00 左右达到全天峰值6.48 μg·cm-2·s-1。乌柳的蒸腾午休现象说明其对环境变化较为敏感,在中午关闭自身气孔减少蒸腾失水以响应高温、低湿、高光强胁迫,表明其对干旱环境有着较好的适应能力。柠条与乌柳类似,清晨即表现出较高的蒸腾强度,在11:00达到全天的次峰值,经过蒸腾午休,于 15:00 达到全天峰值 11.10 μg·cm-2·s-1,之后蒸腾速率迅速下降,表现出良好的自身水分调节机制,但这与齐化龙等[21]测得柠条蒸腾速率在傍晚有一个小幅回升的研究结果有一定差异。
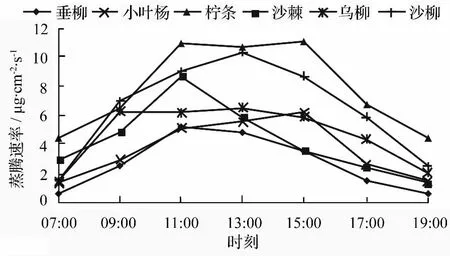
图1 植被恢复树种蒸腾速率日变化
如表1所示,对各树种蒸腾速率特征值采用Duncan方法进行多重比较,结果表明,柠条、沙柳的日均蒸腾速率显著高于垂榆、小叶杨和沙棘(P<0.05),乌柳和柠条之间也存在显著差异(P<0.05);柠条、沙柳的最大蒸腾速率显著高于垂榆和小叶杨(P<0.05),沙棘则与其他树种之间均无显著性差异。综合不同树种日均蒸腾速率及最大蒸腾速率进行系统聚类,结果表明,供试树种可分为两类,其中,柠条、沙柳为高蒸腾速率树种,垂榆、小叶杨、沙棘、乌柳为低蒸腾速率树种。

表1 植被恢复树种蒸腾速率特征值的方差分析 μg·cm-2·s-1
3.2 植物不同方位叶片蒸腾速率日动态
在有关蒸腾的研究中,研究者更注重对植株整体蒸腾特征的分析,而对不同方位叶片蒸腾特征的差异则较少触及,文中对此进行了一定的探析。从图2中可以看出,相同树种不同方位叶片的蒸腾速率日进程曲线基本相同,但也存在一定差异,如柠条和乌柳部分叶片出现了午休现象,而其余方位叶片则表现单峰进程。观察蒸腾速率峰值出现的时间,小叶杨和沙棘各方位叶片保持一致,其余树种则表现出差异性。垂榆西面和北面叶片的蒸腾速率在11:00即达到最大值,早于东面和南面叶片;与之相反,柠条则是东面和南面叶片首先达到蒸腾速率峰值,这表明各树种具有不同的蒸腾耗水适应机制。由供试树种分析,东、南方位叶片峰值时间呈现更大的一致性,应与其受光时间相近且微环境下气温及水分随时间的变化类似相关。从表2可以看出,多数树种南面叶片的日均蒸腾速率最高,其次为西面,东面和北面叶片的日均蒸腾速率则较低,只有垂榆东面叶片的日均蒸腾速率高于南面叶片,不同方位叶片日均蒸腾速率由大到小变化总体趋势为:南、西、东、北。该差异主要是受日照时间、光强影响,南面叶片全天受光,日照时间长、光照强度大、空气湿度较低,导致其蒸腾强度较大,北面叶片则相反。同时,该差异也与不同方位叶片长期适应其环境而产生的自身生理响应相关[22]。

图2 植被恢复树种不同方位叶片蒸腾速率日变化

表2 不同树种不同方位叶片蒸腾速率日均值
3.3 植物瞬时蒸腾量日动态及日蒸腾量
植物通过根系吸收的水分除极少量用于自身生长发育外,大部分被蒸腾作用所消耗[23],因此,植物的蒸腾耗水量直接关系到生态恢复区域的生态需水量。文中对研究区域植被恢复树种蒸腾耗水量进行计算,结果如表3所示,不同树种蒸腾耗水量差异巨大,小叶杨为高大乔木,日蒸腾量可达93.556 kg,而灌木树种沙棘仅为5.866 kg。
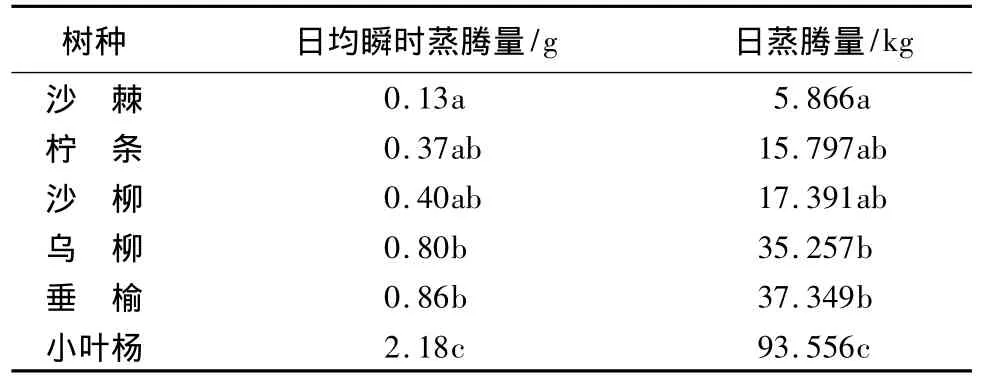
表3 植被恢复树种蒸腾量特征值的方差分析
从图3可以看出,虽然各树种蒸腾速率值相差不大,但是由于不同树种生物量与总叶面积存在较大差异,导致其瞬时蒸腾量差异显著。对各树种蒸腾量特征值采用Duncan方法进行多重比较,结果表明,小叶杨在日均瞬时蒸腾量和日蒸腾量方面均显著高于其他树种(P<0.05);沙棘的蒸腾量特征值也与乌柳、垂榆呈现出显著性差异(P<0.05)。成年的小叶杨由于其较大的生物量及总叶面积,导致其具有明显高于其余树种的蒸腾耗水量,沙棘作为灌木树种,较小的生物量也是其蒸腾耗水量少的主要原因。对植被恢复树种日均瞬时蒸腾量及日蒸腾量进行系统聚类,可将供试树种分为3类,其中,柠条、沙棘、沙柳为低蒸腾耗水树种,垂榆、乌柳为亚高蒸腾耗水树种,小叶杨为高蒸腾耗水树种。20年生乌柳的蒸腾量接近成年的乔木树种垂榆,已经表现出小乔木的特性。柠条、沙柳与沙棘均为灌木树种,前两者蒸腾量高出后者1倍,说明沙棘对干旱环境有更好的适应能力,但考虑供测植物种间生境差异,柠条、沙棘生长于固定沙丘,其土壤水分含量低于丘间低地,植株蒸腾所需水分供应较弱,对其蒸腾量较小也存在一定影响。
4 结论与讨论
供试树种蒸腾速率日动态以单峰型曲线为主,包括垂榆、小叶杨、沙棘、沙柳;柠条和乌柳呈不典型的双峰型曲线,表现出不明显的蒸腾午休现象。植物不同方位叶片蒸腾速率变化趋势基本相同,但也存在一定差异。各树种不同方位叶片蒸腾速率峰值出现的时间不同,东面、南面叶片达到峰值的时间更趋一致。总体上分析,日均蒸腾速率由大到小的变化趋势为:南面叶片、西面叶片、东面叶片、北面叶片。不同树种蒸腾耗水量差异巨大,小叶杨日蒸腾耗水量可达93.556 kg,20年生乌柳对水分的消耗与小乔木垂榆相近,约为小叶杨的40%,灌木树种沙棘、柠条、沙柳耗水量较低。

图3 植被恢复树种瞬时蒸腾量日变化
由共和盆地所处地理位置及气候区域所限,水分将长期成为该地区植被重建的主要限制因子。根据对供试树种的分析,小叶杨蒸腾速率较小,但日蒸腾量最大,不适宜大面积种植;乌柳蒸腾速率较高,多年生乌柳生物量、日蒸腾量也较大,栽植时应考虑其初始间距及空间配置方式,避免高度的种内竞争和对水分的过度消耗;灌木树种柠条、沙柳蒸腾速率较大,但日蒸腾量较低,适合作为该地区的植被恢复树种;沙棘日蒸腾量仅为沙柳的1/3,适宜在该区域广泛种植,作为植被恢复的先锋植物。同时,应考虑在植被恢复过程中采用当地乡土树种,经过长期进化的乡土树种能够对当地的环境变化产生更好的响应。文中研究结果给出了供试树种日均蒸腾速率、最大蒸腾速率及日蒸腾量等指标,该区域可结合当地水分状况、景观格局,合理选取上述指标,以更好地达到生态恢复和荒漠治理的目的。
[1] 郭连云,丁生祥,汪青春.气候变化与人类活动对共和盆地生态环境的影响[J].干旱地区农业研究,2009,27(1):219-226.
[2] 杨芳.气候变化对青海东部降水和径流的影响[J].人民黄河,2011,33(7):37-41.
[3] 周陈超,贾绍凤,燕华云,等.近50a以来青海省水资源变化趋势分析[J].冰川冻土,2005,27(3):432-437.
[4] 姜凤歧,曹成有,曾德息.科尔沁沙地生态系统退化与恢复[M].北京:中国林业出版社,2002.
[5] 丁越岿,杨劼,张仲平,等.毛乌素沙地主要建群植物蒸腾耗水特性的研究[J].中国草地学报,2011,33(4):37-43.
[6] 牛丽丽,张学培,曹奇光.植物蒸腾耗水研究[J].水土保持研究,2007,14(2):158-160.
[7] 张劲松,孟平,尹昌君,等.农林复合系统的水分生态特征研究述评[J].世界林业研究,2003,16(1):10-14.
[8] Andreas R,Sandra K,Sten G.The Climate-Species-Matrix to select tree species for urban habitats considering climate change[J].Urban Forestry & Urban Greening,2009,8:295-308.
[9] Wang H,Ouyang Z Y,Chen W P.Water heat and airborne pollutants effects on transpiration of urban trees[J].Environmental Pollution,2011,159:2127-2137.
[10] 张建国,李吉跃,沈国舫.树木耐早特性及其机理研究[M].北京:中国林业出版社,2000.
[11] 李永华,罗天祥,卢琦,等.青海省沙珠玉治沙站17种主要植物叶性因子的比较[J].生态学报,2005,25(5):994-999.
[12] 石蒙沂.共和盆地柽柳沙地造林技术研究[J].干旱区研究,1999,16(4):61-64.
[13] 魏占雄.高寒沙区生态恢复对植物物种多样性的影响[J].草业与畜牧,2009(7):36-39.
[14] 屈艳萍,康绍忠,张晓涛,等.植物蒸发蒸腾量测定方法述评[J].水利水电科技进展,2006,26(3):72-77.
[15] 苏建平,康博文.我国树木蒸腾耗水研究进展[J].水土保持研究,2004,11(2):177-179.
[16] 巨关升,刘奉觉,郑世锴.选择树木蒸腾耗水测定方法的研究[J].林业科技通讯,1998(10):12-14.
[17] 杨洪晓,卢琦,吴波,等.高寒沙区植被人工修复与种子植物物种多样性的变化[J].林业科学,2004,40(5):45-49.
[18] 刘君梅,王学全,刘丽颖,等.高寒沙区植被恢复过程中表层土壤因子的变化规律[J].东北林业大学学报,2011,39(8):47-49.
[19] 董学军,杨宝珍,郭柯,等.几种沙生植物水分生理生态特征的研究[J].植物生态学报,1994,18(1):86-94.
[20] 段爱国,张建国,张守攻,等.干热河谷主要植被恢复树种蒸腾作用[J].生态学报,2009,29(12):6691-6701.
[21] 齐化龙,张维江.毛乌素沙地沙生植物蒸腾规律探讨[J].农业科学研究,2008,29(1):61-65.
[22] 李黛,谈锋.非均衡环境下海桐和日本珊瑚树水分生理的适应性研究[J].遵义师范学院学报,2005,7(2):54-56.
[23] 武维华.植物生理学[M].北京:科学出版社,2003:53-66.
猜你喜欢
现代园艺(2022年16期)2022-09-06
现代农村科技(2021年5期)2021-12-07
乡村科技(2021年16期)2021-10-21
西南农业学报(2021年7期)2021-08-23
节水灌溉(2021年5期)2021-05-26
中国煤炭工业(2020年1期)2020-12-28
中国农业科技导报(2020年12期)2020-03-15
当代陕西(2019年7期)2019-04-25
散文诗世界(2016年11期)2016-11-16
中国塑料(2016年12期)2016-06-15
