
大兴安岭低质林不同林型主要树种冠层分析与比较1)
2013-08-08 07:21:52董希斌宋启亮曾翔亮
东北林业大学学报 2013年9期
崔 莉 董希斌 宋启亮 曾翔亮
(东北林业大学,哈尔滨,150040)
低质林是由于强烈的自然灾害和人为因素干扰破坏,林分的组成成分发展缓慢[1],甚至逆向发展,造成林分郁闭度降低、林分产量下降、林地土壤侵蚀严重,整个森林生态系统的自我恢复能力很低,该区域的森林生态效益和经济价值降低。目前,低质林改造研究主要集中在改造方式对林分的水土保持、土壤温湿度、土壤呼吸[2]、土壤理化性质[3]、生物多样性和生物量等方面[4],通过这些研究确定合理的改造方式,提高低质林的林分综合质量。
冠层是植物群体地上部分的绿色覆盖层,包括植物的叶、茎、枝条、花和果实等器官,冠层的大小、形状、方位和在冠层中的上下位置的分布情况。冠层结构是判断群落外观的可视化指标,冠层结构对植被截获太阳辐射的程度、风速、空气温湿度、土壤蒸发量、土壤温度等各个不同方面都有影响,能够调节植物与环境的相互作用,同时对动植物生存和生长都有一定的影响[5],冠层结构指数可以直接反映植被的生长能力,冠层研究有助于探究该地区的环境因子[6]。
1 试验地区概况
试验区地处黑龙江省大兴安岭地区最南部,属于低山丘陵地带,地理坐标为东经124°23'47.8″~124°24'35.1″,北纬 50°34'9.17″~ 50°34'32.0″。位于大兴安岭加格达奇翠峰林场174林班,在大兴安岭山脉的东南坡,属于限伐区,该地区地势平缓,坡度在10°左右,海拔高度为370~420 m,立地条件好。土壤以暗棕壤和棕色针叶林土为主;属于寒温带大陆季风气候,冬长夏短,且冬季气候寒冷,年平均气温-1.3℃,最高气温37.3℃,最低气温-45.4 ℃,年平均降水量为 494.8 mm,多发生在6、7、8 月份。
在大兴安岭林区加格达奇林业局翠峰林场174林班选取3种典型低质林林分类型进行试验,3种类型低质林分别为山杨(davidiana)低质林、白桦(Betula platyphylla Suk)低质林和蒙古栎(Xylosma racemosum)低质林,3种林型的立地条件和林分概况见表1。

表1 3种类型低质林立地条件和林分概况
2 材料与方法
Winscanopy植物冠层分析系统主要包括:Winscanopy分析软件、XLScanopy数据处理软件、高分辨率专业数码相机及180°鱼眼镜头等。Winscanopy通过由数码相机和鱼眼镜头拍摄的半球图像实现分析。选择树木进行实地拍摄,使用鱼眼镜头拍摄所要研究的植被冠层,获得半球状的图像,再利用Winscanopy软件对图像进行处理,获得有关植被冠层的相关数据后对太阳光直射透过的系数进行计算。
于2012年8月中旬在加格达奇翠峰林场174林班的山杨低质林、白桦低质林和蒙古栎低质林分别随机选取50棵树,用GPS分别测得每棵树木的所在地点的经纬度和海拔高度,用数据采集装置Mini-O-Mount 7MP进行实地的试验,找准正北方向并调平仪器,测量仪器镜头离地距离,从3~4个不同方向进行观测,采集图像。
使用冠层分析仪Winscanopy处理采集到的图像得到初步的试验数据,再用XLScanopy对数据进行校正等预处理,最后导入Excel和SPSS对数据进行计算处理。
3 结果与分析
3.1 林隙分数与开度
林隙分数是指图像中像素等级作为开放天空(不包括植被阻隔的)所占图像(在两个空间间隔)中天空网格区域的指数;开度是林隙分数经过补偿计算剔除了植被阻隔的影响得出的实际冠层林隙分数[7],二者都是体现冠层透光率的指标。
通过数据分析得出3个树种林隙分数和开度的拟和曲线(图1)。在大兴安岭低质林中,运用冠层分析仪测得的林隙分数和开度。山杨林、白桦林和蒙古栎林中林隙分数分别为10.69% ~1.79%、9.34% ~22.85%和 5.72% ~14.27%,平均值分别为 14.98%、14.07%和9.63%;开度分别在11.32% ~26.52%、9.91% ~ 24.44% 和 5.85% ~ 15.12%,平均值分别为15.94%、15.86%和9.98%。测定结果没有受到不同树种树木生长特性不同的影响,山杨林、白桦林和蒙古栎林中试验结果说明林隙分数和开度的相关性极强,且两组值差异不显著,开度随着林隙分数的增加呈明显上升趋势,说明3种林型下林隙分数与开度的关系都呈显著正相关关系,林隙分数和开度相关性越显著,枝叶阻隔对林隙分数的影响程度越小。山杨林中林隙分数和开度在3个树种中最大,白桦林次之,蒙古栎林最小。由显著性水平R2值证明,蒙古栎林中枝叶阻隔对林隙分数的影响程度最小。
3.2 林隙分数与叶面积指数
叶面积指数(LA,I)是指绿叶的总面积占单位水平种植面积的比值。通过叶面积指数可以对植物进行生物量和地球生态系统能量交换特性进行定量分析,叶面积指数与树冠的光合作用、蒸腾作用以及生产力等方面密切相关,LA,I决定了陆地表面植被的生产力,同时对地表和大气之间的相互作用也有一定的影响。尽管它的定义非常简单,但是要准确地测量出叶面积指数也比较困难[8-9]。叶面积指数是植被生态系统的一个重要结构参数。植物叶片影响着林木冠层内的许多生物化学过程,在生态过程、大气生态系统的交互作用以及全球变化等研究中都需要用到叶面积指数这一参数[10-13]。

图1 3个树种林隙分数和开度的拟合曲线
以前的研究者测量叶面积指数采用直接测量法或者间接测量法等[14-15]。冠层分析仪能够得出4种方法测量所得叶面积指数的值,高登涛等[16]对实测的LA,I结果与Winscanopy的分析结果进行相关性检验,发现实测结果与 LA,I(2000)-Log方法的相关性最好,本试验中采用的叶面积指数取分析结果中LA,I(2000)-Log的值。
经过数据分析得出3个树种林隙分数与叶面积指数的拟合曲线(图2)。可以看出,3个树种林隙分数与叶面积指数呈现出负相关的关系,随着林隙分数的增大,林中透光率增加,冠层下方光照增加,相应的单位面积上叶片的覆盖率较小,所以叶面积指数随之减小。山杨林、白桦林和蒙古栎林中叶面积指数值分别为0.97 ~2.57、1.02 ~3.23 和2.74 ~5.38,平均值分别为 1.92、2.03 和 3.99,试验得出该地区的叶面积指数蒙古栎林较白桦林和山杨林都大。

图2 3个树种林隙分数与叶面积指数的拟合曲线
3.3 林隙分数与总定点因子
总定点因子是定量表示单位时间内透过冠层光的与光照入射辐射有关的数据。也可定义为透过冠层接收到的日平均辐射占冠层上方入射光辐射的比例,也即透光率[7]。
由图3可看出,总定点因子随林隙分数的增加均呈直线上升,说明在冠层上方接受到的日平均光合辐射总量基本相同的情况下,阳光随林隙的增加,透过冠层到达冠下的总光合有效辐射平均通量密度(下文简称冠下总光合辐射)明显上升,即林隙分数增大,林中透光率增加,透过冠层的光辐射较强,冠层下方接收的日平均辐射量增加,总定点因子随之增加。3种林分相比较,由大到小的顺序为山杨林(0.298 4)、白桦林(0.252 1)、蒙古栎林(0.177 2)。

图3 3个树种林隙分数与总定点因子的拟合曲线
3.4 叶面积指数与冠下总光合辐射
植物的光合作用面积与叶面积指数相关,通常用叶面积指数来表示光和面积,总定点因子表明透光率的大小,可以用来表示阳光透过冠层到达冠下的能力,也能够表示植被冠层获得光能力的强弱,冠层结构决定太阳辐射在冠层内的分布,由太阳辐射和冠层结构可以计算出辐射在冠层内部的分布[17-19],冠层内的有效光合辐射分布对冠层光合作用有着重要的影响,同时温度、湿度、风速和土壤养分等因子对光合作用也有很大的影响,这些影响也与冠层结构相关[20-21]。
由图4可看出,叶面积指数和冠下总光合辐射呈显著负相关关系。随着叶面积指数增大,单位面积的叶片覆盖率增加,植被对阳光的获截能力提升,透过树冠到达冠层下方的辐射量减少,从而导致冠下总光合辐射减小。试验得出山杨林、白桦林和蒙古栎林的叶面积指数分别为0.97 ~2.57、1.02 ~3.23和2.74 ~5.38,平均值分别为1.92、2.03和3.99;冠下总光合辐射量分别为3.45 ~11.57、2.11 ~6.53、3.61 ~ 6.53 mol·m-2·d-1,平均值分别为5.62、4.77 和4.15 mol·m-2·d-1。结果表明,大兴安岭地区蒙古栎林的光获截能力最强,山杨林最弱。
3.5 叶面积指数与总定点因子
通过对不同的树种叶面积指数和总定点因子测量结果进行相关性分析,得到3个树种叶面积指数与总定点因子的拟合曲线(图5)。得出不同树种的叶面积指数与总定点因子的相关性变化程度不完全相同,随着叶面积指数的增大,总定点因子都随之减小,皆呈现明显的负相关性。树冠透光率减少时,说明单位面积叶片面积所占比例较大,总定点因子减小。对于同一层面,叶面积指数越大,树冠的光能利用率较高,此时总定点因子较小。

图4 3个树种叶面积指数与冠下总光合辐射拟合曲线

图5 3个树种总定点因子与叶面积指数拟合曲线
山杨林、白桦林和蒙古栎林中总定点因子值分别为0.160 5 ~0.496 2、0.131 0 ~0.274 8、0.171 2 ~0.256 8,平均值分别为0.298 4、0.252 1 和0.177 2,从定点因子这一因素验证了3种林分的光获截能力由大到小的排序为蒙古栎林、白桦林、山杨林。
3.6 3个树种光获截能力的分析比较
冠层光合有效辐射平均通量密度是评价冠层光截获能力最重要的指标,冠层上方总的光合有效辐射平均通量密度与冠层下方的总光合有效辐射平均通量密度之差,即冠层的光截获密度[22]。在同一试验地中,冠层上方的光照辐射强度相同,冠层上方总光合有效辐射平均通量密度基本相同,冠层的光获截能力直接影响到冠层下方的总光合辐射,因此,冠下总光合辐射大小反映冠层的光获截能力强弱,由于不同树种冠层的差异很大,对阳光的截获能力也有很大差异。通过分析得出3个树种的总定点因子与冠下总光合辐射的拟合曲线(图6)。从图6可看出,总定点因子与冠层下方总辐射通量平均密度相关性极显著,呈现出显著的正相关关系。山杨林、白桦林和蒙古栎林的冠下总光合辐射分别为 3.45 ~11.57、2.11 ~6.53和 3.61 ~6.53 mol·m-2·d-1,平均值分别为 5.62、4.77 和 4.15 mol·m-2·d-1。对 3 个树种的冠下光合辐射进行比较(表2),结果表明,林木冠下总光合辐射主要来自于冠下直接光合辐射,同时表2中的数据再次验证了蒙古栎林冠层的光获截能力最强。
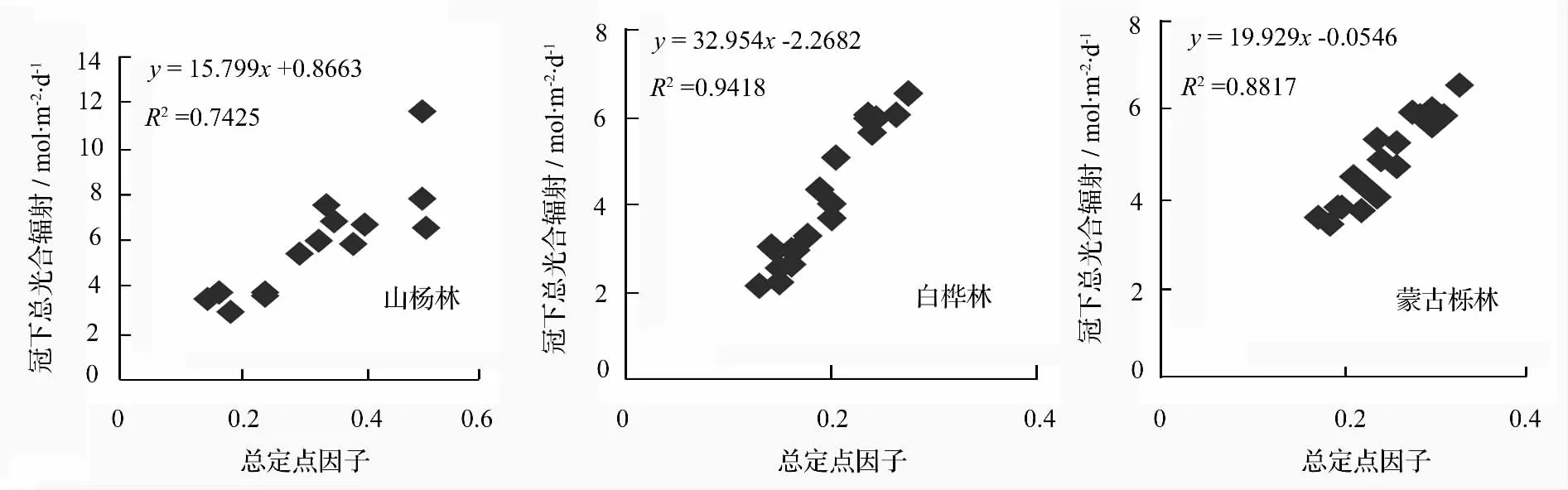
图6 3个树种总定点因子和冠下总辐射拟合曲线

表2 不同树种的冠下光合辐射比较 mol·m-2·d-1
4 结论与讨论
通过对大兴安岭地区加格达奇翠峰林场174林班的山杨(Populus davidiana Dode)低质林、白桦(Betula platyphylla Suk)低质林和蒙古栎(Xylosma racemosum)低质林3种林分进行冠层分析,得到试验结果如下:
大兴安岭地区山杨林、白桦林和蒙古栎林的冠层结构指标测定结果分别为:林隙分数14.97%、14.07%、9.63%,开度 15.94%、15.86%、9.98%,叶面积指数 1.92、2.03、3.99,总定点因子 0.2984、0.2521、0.1772,生长季节冠层下方总光合有效辐射平均通量密度 5.62、4.77 和 4.15 mol·m-2·d-1。
对于不同的树种,冠层特征指标林隙分数、冠层的光合有效辐射平均通量密度和叶面积指数都有显著的相关性。试验得出:林隙分数与开度、总定点因子都呈现显著正相关,叶面积指数LA,I与林隙分数、总定点因子及冠下总光合辐射皆为负相关,总定点因子与冠下总光合辐射呈正相关。
对大兴安岭地区低质林中3个树种的冠层特性指标进行分析,该地区蒙古栎林的冠层光获截能力最强,白桦林次之,山杨林最弱。3种林分由于离地因子的差异,由此可能导致入射光的不同,高荣孚[23]研究发现植物群体内部的辐射状况不仅仅取决于入射光的数量,而且与植物个体和群体的结构息息相关。不同的树种叶片分布的差异直接影响到自身光合作用和呼吸作用的强弱,过密的叶片分布会造成相互遮光,从而降低了植被吸收光能的效率,使叶片的平均光合效率降低,同时又增加了植被光合产物的消耗[24],对植物的生长不利。不同的立地条件土壤的理化性质也会有很大的差异,高健等[25]研究发现地形对杨树的光合作用有很大的影响,这些因素都有可能造成大兴安岭地区不同林型树种的冠层参数的差异,对于主要影响因素,还有待进一步的试验和论证。
[1] 杨学春,董希斌,姜帆.黑龙江省伊春林区低质林林分评定[J].东北林业大学学报,2009,37(10):10-15.
[2] 郭辉.采伐对小兴安岭低质林土壤和枯落物功能影响[D].哈尔滨:东北林业大学,2010.
[3] 张泱,宋启亮,董希斌.不同采伐强度改造对小兴安岭低质林土壤理化性质的影响[J].东北林业大学学报,2011,39(11):22-24.
[4] 李勇,刘继明,董希斌,等.大兴安岭林区低质林成因及改造方式[J].东北林业大学学报,2012,40(8):105-107.
[5] 刘晓东,朱春全,雷静品,等.杨树人工林冠层光合辐射分布的研究[J].林业科学,2000,36(3):2-7.
[6] 孙婧,王刚,孟艳琼,等.不同环境下黄山杜鹃光合特性及其与主要环境因子的关系[J].东北林业大学学报,2013,41(3):9-12.
[7] Jelaska S D.Analysis of canopy closure in the Dinaric silver firbeech forests(Omphalodo-Fagetum)in Croatia using hemispherical photography[J].Hacquetia,2004,3(2):43-49.
[8] Chen J M,Cihlar J.Retrieving leaf area index of boreal conifer forests using Landsat TM images[J].Remote sensing of environment,1996,55(2):153-162.
[9] Chason J W,Baldocchi D D,Huston M A.A comparison of direct and indirect methods for estimating forest canopy leaf area[J].Agricultural and Forest Meteorology,1991,57(1):107-128.
[10] Pierce L L,Running S W.Rapid estimation of coniferous forest leaf area index using a portable integrating radiometer[J].Ecology,1988,69(6):1762-1767.
[11] Gholz H L,Vogel S A,Cropper Jr W P,et al.Dynamics of canopy structure and light interception in Pinus elliottii stands,north Florida[J].Ecological Monograph,1991,61(1):33-51.
[12] 田庆久,闵祥军.植被指数研究进展[J].地球科学进展,1998,13(4):327-333.
[13] 薛利红,曹卫星,罗卫红,等.光谱植被指数与水稻叶面积指数相关性的研究[J].植物生态学报,2004,28(1):47-52.
[14] Frazer G W,Fournier R A,Trofymow J A,et al.A comparison of digital and film fisheye photography for analysis of forest canopy structure and gaplight transmission[J].Agricultural and Forest Meteorology,2001,109(4):249-263.
[15] van Gardingen pr,Jackson G E,Hernandez-Daumas S,et al.Leaf area index estimates obtained for clumped canopies using hemispherical photography[J].Agricultural and Forest Meteorology,1999,94(3):243-257.
[16] 高登涛,韩明玉,李丙智,等.冠层分析仪在苹果树冠结构光学特性方面的研究[J].西北农业学报,2006,15(3):166-170.
[17] 张小全,徐德应,赵茂盛.林冠结构、辐射传输与冠层光合作用研究综述[J].林业科学研究,1999,12(4):411-421.
[18] 牛文元,周允华,张翼.农田生态系统能量物质交换[M].北京:气象出版社,1987.
[19] 孟平.农林复合生态系统研究[M].北京:科学出版社,2004.
[20] Farquhar G D,von Caemmerer S,Berry J A.A biochemical model of photosynthetic CO2assimilation in leaves of C3species[J].Planta,1980,149(1):78-90.
[21] Leaning R.A critical appraisal of a combined stomata-photosynthesis model for C3plants[J].Plant Cell Environ,1995,18(4):339-355.
[22] 刘立鑫.应用冠层分析仪对天然次生林冠层结构及光照分布的研究[D].哈尔滨:东北林业大学,2009.
[23] 高荣孚.杨树光合作用研究进展[J].北京林学院学报,1981,3(1):56-62.
[24] 温志宏,黄敏仁.美洲黑杨冠层光截获特性的遗传学研究[J].南京林业大学学报,1992,16(3):11-17.
[25] 高健,吴泽民,彭镇华.滩地杨树光合作用生理生态的研究[J].林业科学研究,2000,13(2):147-152.
猜你喜欢
中国农业信息(2022年1期)2022-05-25 13:31:46
农业机械学报(2021年11期)2021-12-07 05:36:44
湖南农业大学学报(自然科学版)(2021年3期)2021-07-02 01:48:06
大气科学(2021年1期)2021-04-16 07:34:18
新疆农业科学(2020年1期)2020-02-14 03:22:46
海峡姐妹(2019年8期)2019-09-03 01:00:56
海峡姐妹(2018年11期)2018-12-19 05:18:28
东北林业大学学报(2017年10期)2017-10-24 11:50:18
林业科技(2017年4期)2017-08-10 06:02:43
农业环境科学学报(2017年2期)2017-03-20 14:57:37
