
人乳腺癌上皮细胞miR-217对EZH2表达的调控作用
2013-07-05 09:39王璐璐刘萃萃赵卫卫王玉平孙振亮
山东医药 2013年43期
王璐璐,刘萃萃,郭 腾,陶 然,彭 攸,赵卫卫,王玉平,孙振亮,冯 景
(1苏州大学医学部,江苏苏州215000;2上海市奉贤区中心医院)
乳腺癌的发生发展是一个多因素、多阶段的过程,其中组蛋白甲基化及DNA甲基化等发挥了重要作用[1,2]。homolog2 zeste 基因增强子(EZH2)定位于染色体7q35-6,是PcG基因家族的重要成员[3],通过诱导组蛋白乙酰化和甲基化等表观遗传的发生来调节基因的表达[4]。EZH2高表达于多种肿瘤,如乳腺癌[5]。在前期研究中[6],我们利用经典分子生物学方法,结合慢病毒lenti-miRNAs文库筛选出了包涵miR-217在内的17个靶向EZH2的miRNAs。2012年3月21日~12月10日,我们通过RT-PCR、Western blot和双荧光素酶报告基因等一系列实验方法,进一步研究乳腺上皮细胞系中miR-217与EZH2之间的作用机制,为miR-217作为乳腺癌潜在治疗靶点的体外研究提供实验依据。
1 材料与方法
1.1 材料 人正常乳腺上皮细胞系HBL-100,乳腺癌上皮细胞系 MCF-7、MDA-MB-231,人胚肾细胞293T。DMEM高糖培养基。0.25%胰蛋白酶、青—链霉素(10 000 U/mL)。胎牛血清。TRIzol。逆转录试剂盒。实时定量PCR用 2×Mix、Syber Green染料、Mg2+。EZH2,18s、U6引物。5×蛋白裂解液。一抗EZH2。β-actin,二抗羊抗鼠。免疫印迹凝胶配置试剂。免疫印迹配胶、电泳、电转装置。A、B显影液。转染试剂Lipofecter C0516。miR-217检测试剂盒,miR-217模拟物(miR-217 minics)、miR-217抑制剂(miR-217 inhibitor)转染试剂盒。双荧光素酶检测试剂盒,psiCHECKTM-2双荧光素酶载体。
1.2 实验方法
1.2.1 HBL-100、MCF-7、MDA-MB-231 细胞中EZH2、miR-217基因检测 用TRIzol提取对数生长期的单层培养细胞总RNA,用逆转录试剂盒将总RNA逆转录合成cDNA,进行RT-PCR。EZH2引物序列:上游为5'-TTCGGTAAATCCAAACTGCTAT-3',下游为5'-GCCTGGCTGTATCTGTAATCAA-3';扩增产物126 bp;反应条件:94℃预变性3 min,94℃变性30 s,55 ℃ 退火 30 s,72 ℃ 延伸 30 s,共 35 个循环。miR-217引物序列:上游为 5'-TACTCAACTCACTACTGCATCAGGA-3',下游为 5'-TATGGTTGTTCTGCTCTCTGTGTC-3';扩增产物84 bp;反应条件:95℃预变性3 min,95℃变性12 s,60℃退火、延伸20 s,共40个循环。实验重复2次。由PCR反应曲线得到域值循环数(Ct),以基因18s或U6作为内参照,计算基因相对表达量。
1.2.2 HBL-100、MCF-7、MDA-MB-231 细胞中EZH2蛋白检测 采用Western blot法。分别将收集的细胞加入适量蛋白裂解液,摇床上冰浴裂解15 min,加入适量蛋白loading buffer,100℃干浴锅煮10 min,室温冷却后4℃保存。每泳道20μL蛋白样品进行十二烷基硫酸钠—聚丙烯酰胺凝胶电泳,转硝酸纤维素膜,鼠抗人EZH2(1∶300)过夜,二抗羊抗鼠(1∶300)室温孵育3 h,暗室中A、B液显色,显影,定影。以β-actin作为参照。蛋白表达量采用Launch Sensiansys软件进行灰度分析。
1.2.3 microRNAs的靶基因预测 采用 DIANA-microT、miRanda、PicTar、Target Scans 软件共同预测miR-217的靶基因。
1.2.4 miR-217过表达与抑制实验 将实验细胞以2×105/孔接种于6孔培养板。将10 nmol/L的miR-217 minics(实验组)及 miR-217 minics control(对照组)分别转染到低表达miR-217的MDA-MB-231细胞中。将50 nmol/L的miR-217 inhibitor(实验组)及miR-217 inhibitor control(对照组)分别转染到高表达 miR-217的HBL-100细胞中。37℃的CO2培养箱孵育48 h后收集细胞。观察miR-217、EZH2蛋白的表达。
1.2.5 双荧光素酶检测实验 将含有miR-217识别位点的312 bp的EZH2 3'UTR(EZH2-UTR-WT)进行PCR扩增,并将PCR产物亚克隆于psiCHECKTM-2双荧光素酶载体上。海肾荧光素酶位于插入片段的上游,萤火虫荧光素酶位于插入片段的下游。应用NCBI BLAST比对,miR-217与EZH2 3'UTR有8个连续的互补配对碱基(图1),将这8个碱基突变掉,构建含miR-217突变位点的psiCHECKTM-2载体,即EZH2-UTR-MUT。利用转染试剂Lipofecter C0516将 1μg的质粒和 50 nmol/L的 miR-217 minics转染到293T细胞中。37℃的CO2培养箱孵育48 h后裂解细胞,进行荧光素酶活性检测。相对荧光素酶活性为海肾荧光素酶活性/萤火虫荧光素酶活性。

图1 miR-217与EZH2 3'UTR有8个连续的互补配对碱基
1.3 统计学方法 采用SPSS17.0统计软件。计量资料以¯x±s表示,比较用t检验和单因素方差分析(ANOVA)。P≤0.05为差异有统计学意义。
2 结果
2.1 乳腺上皮细胞系中miR-217的转录与表达与HBL-100比较,miR-217在 MCF-7和 MDA-MB-231中显著低表达(图2)。

图2 三种细胞中miR-217的表达
2.2 HBL-100比较,EZH2 mRNA在 MCF-7中高表达、MDA-MB-231中显著高表达(图3)。HBL-100中EZH2蛋白低表达,MCF-7、MDA-MB-231中EZH2蛋白高表达(图4)。

图3 三种细胞中EZH2 mRNA的表达

图5 实验组和对照组MDA-MB-231细胞miR-217的表达
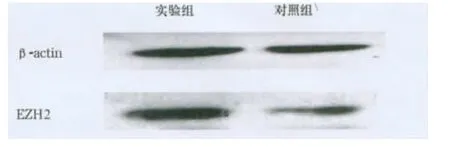
图6 实验组和对照组MDA-MB-231细胞EZH2蛋白的表达

图4 三种细胞中EZH2蛋白的表达
2.3 过表达或抑制miR-217对EZH2表达的影响PicTar、miRanda、Target Scans软件都预测出 EZH2是miR-217的靶基因。MDA-MB-231细胞转染miR-217 minics后,实验组miR-217表达量是对照组的21.67倍(图5);与对照组相比,实验组EZH2蛋白表达量下降了71.96%(图6)。HBL-100细胞转染miR-217 inhibitor后,与对照组相比,实验组miR-217表达量下降了85.65%(图7),EZH2蛋白表达量增加了 179.63%(图 8)。
2.4 miR-217对EZH2的靶向调控作用 双荧光素酶报告实验结果显示,293T细胞分别转染EZH2-UTR-WT和 EZH2-UTR-MUT载体48 h后,野生型EZH2-UTR-WT的相对荧光素酶活性是3.525,突变型EZH2-UTR-MUT的相对荧光素酶活性是5.365,两者相比,P <0.05。

图7 实验组和对照组HBL-100细胞miR-217的表达

图8 实验组和对照组HBL-100细胞EZH2蛋白的表达
3 讨论
研究表明,miRNAs通过与EZH2的3'UTR结合来调节EZH2表达,如 miR-26a、miR-98、miR-101、miR-124、miR-138和miR-214可分别在鼻咽癌、恶性胶质瘤、肝癌、头颈部鳞状细胞癌和神经母细胞瘤中通过靶向EZH2而抑制EZH2的表达[7~12]。
miR-217作为miRNAs家族中的一员,是一种多功能的miRNA。例如,miR-217可以通过靶向作用于KRAS抑制胰腺导管癌的发生[13];在大鼠肾小球系膜细胞(MC)中 miR-216a协同 miR-217靶向PTEN 可促使 TGF-β 激活 AKt[14];在人内皮细胞中,随着年龄的增长miR-217逐渐增多,从而抑制SirT1的表达,诱导细胞的早期衰亡[15];在人胚肾293细胞中共转染miR-217和miR-377,HO-1蛋白表达下降,HO-1酶活性减低;反之,将miR-217和miR-377同时干扰掉,HO-1蛋白的表达上升[16];miR-217与miR-196a可作为判定是否患有胰腺导管癌或慢性胰腺炎的一个分子生物学标志等[17]。但在乳腺上皮细胞中,miR-217与EZH2的作用机制尚不清楚。本研究结果显示,在正常乳腺上皮细胞系HBL-100中,EZH2低表达,miR-217高表达;在乳腺癌上皮细胞系MCF-7和 MDA-MB-231中,EZH2高表达,而miR-217低表达;此结果与Tan等[18]报道相符。瞬时转染miR-217 minics和miR-217 inhibitor,过表达和抑制miR-217,结果显示过表达miR-217时EZH2蛋白表达下降,抑制miR-217时EZH2蛋白表达上升。说明miR-217可以下调EZH2的表达。利用293T细胞作为双荧光素酶报告实验的工具细胞,分别转染EZH2-UTR-WT及EZH2-UTR-MUT载体质粒,48 h后进行双荧光检测,结果显示两者的相对荧光素酶活性差异有统计学意义。说明miR-217通过直接作用于EZH2 3'UTR下调EZH2表达。
总之,本研究结果表明,在乳腺癌上皮细胞中miR-217对EZH2的表达具有调控作用,且miR-217通过直接作用于EZH2 3'UTR调控EZH2表达。这为miR-217可能作为乳腺癌潜在治疗靶点的体外实验研究提供了依据。
[1]Alford SH,Toy K,Merajver SD,et al.Increased risk for distant metastasis in patients with familial early-stage breast cancer and high EZH2 expression[J].Breast Cancer Res Treat,2012,132(2):429-437.
[2]Shi JF,Li XJ,Si XX,et al.ERα positively regulated DNMT1 expression by binding to the gene promoter region in human breast cancer MCF-7 cells[J].Biochem Biophysi Res Commun,2012,427(1):47-53.
[3]Vire E,Brenner C,Deplus R,et al.The Polycomb group protein EZH2 directly controls DNA methylation[J].Nature,2006,439(7078):871-874.
[4]Saramaki OR,Tammela TL,Martikainen PM,et al.The gene for polycomb group protein enhancer of zeste homolog 2(EZH2)is amplified in late-stage prostate cancer[J].Genes Chromosomes Cancer,2006,45(7):639-645.
[5]Kleer CG,Cao Q,Varambally S,et al.EZH2 is a marker of aggressive breast cancer and promotes neoplastic transformation of breast epithelial cells[J].Proc Nati Acad Sci Am,2003,100(20):11606-11611.
[6]彭攸,赵卫卫,李晓娇,等.人EZH2基因编码区及3'非翻译区融合蛋白重组慢病毒表达载体构建及其在293T细胞中的表达[J].中华临床医师杂志,2012,6(24):8151-8155.
[7]Lu J,He ML,Wang L,et al.MiR-26a inhibits cell growth and tumorigenesis of nasopharyngeal carcinoma through repression of EZH2[J].Cancer Res,2011,71(1):225-233.
[8]Alajez NM,Shi W,Hui AB,et al.Enhancer of zeste homolog 2(EZH2)is overexpressed in recurrent nasopharyngeal carcinoma and is regulated by miR-26a,miR-101,and miR-98[J].Cell Death Dis,2010,1(10):85.
[9]Cao P,Deng Z,Wan M,et al.MicroRNA-101 negatively regulates EZH2 and its expression is modulated by androgen receptor and HIF-1alpha/HIF-1beta[J].Mol Cancer,2010,9(1):108-120.
[10]Zheng F,Liao YJ,Cai MY,et al.The putative tumour suppressor microRNA-124 modulates hepatocellular carcinoma cell aggressiveness by repressing ROCK2 and EZH2[J].Gut,2012,61(2):278-289.
[11]Kisliouk T,Yosefi S,Meiri N.MiR-138 inhibits EZH2 methyltransferase expression and methylation of histone H3 at lysine 27,and affects thermotolerance acquisition[J].Eur JNeurosci,2011,33(2):224-235.
[12]Derfoul A,Juan AH,Difilippantonio MJ,et al.Decreased microRNA-214 levels in breast cancer cells coincides with increased cell proliferation,invasion and accumulation of the polycomb EZH2 methyltransferase[J].Carcinogenesis,2011,32(11):1607-1614.
[13]Zhao WG,Yu SN,Lu ZH,et al.The miR-217 microRNA functions as a potential tumor suppressor in pancreatic ductal adenocarcinoma by targeting KRAS[J].Carcinogenesis,2010,31(10):1726-1733.
[14]Kato M,Putta S,Wang M,et al.TGF-β activates Akt kinase via a microRNA-dependent amplifying circuit targeting PTEN[J].Nat Cell Biol,2009,11(7):881-889.
[15]Rossella M,Viviana C,Marina C,et al.MicroRNA 217 modulates endothelial cell senescence via silent Information regulator 1[J].Circulation,2009,120(15):1524-1532.
[16]Joan DB,Chunseng C,Julia N,et al.Regulation of heme oxygenase-1 protein expression by miR-377 in combination with miR-217[J].JBiol Chem,2011,286(5):3194-3202.
[17]Anna ES,Martina D,Hayward SE,et al.Analysis of microRNAs in pancreatic fine-needle aspirates can classify benign and malignant tissues[J].Clin Chem,2008,54(10):1716-1724.
[18]Tan J,Yang X,Zhuang L,et al.Pharmacologic disruption of polycomb-repressive complex 2-mediated gene repression selectively induces apoptosis in cancer cells[J].Genes Dev,2007,21(9):1050-1063.
猜你喜欢
中老年保健(2022年6期)2022-08-19
食品安全导刊(2021年21期)2021-08-30
天津医科大学学报(2021年4期)2021-08-21
中日友好医院学报(2021年1期)2021-04-14
山东医药(2020年9期)2020-05-20
中国生殖健康(2019年2期)2019-08-23
天津医科大学学报(2019年6期)2019-08-13
中国生殖健康(2019年6期)2019-01-06
祝您健康(2018年5期)2018-05-16
现代检验医学杂志(2016年2期)2016-11-14
