
利用β-纤维蛋白原第7内含子分析三种鹤的亲缘关系
2013-07-04 05:45:14陈红李鹏
黑龙江八一农垦大学学报 2013年3期
陈红,李鹏
(黑龙江八一农垦大学动物科技学院,大庆 163319)
鹤类属于鸟纲、鹤形目、鹤科,是世界上珍稀的涉禽鸟类[1]。由于鹤类种群小而分散,繁殖速度缓慢,对狩猎压力和化学污染反应脆弱,从而造成鹤类数量急剧锐减,全世界现存15种,已有7种被列为濒危动物[2]。而我国现存鹤类仅存9种[1]。全面深入的了解鹤类系统发生和基因组的进化将为鹤类的保护工作打开全新的局面,对鹤类的保护和研究具有重要意义。但有关鹤类种群遗传多样性的研究鲜有报道。
与RAPD[3]技术一样,单核苷酸多态性分析是构建系统发育树的重要方法[4]。细胞核基因的内含子是普遍存在的,并且相邻的外显子的核苷酸序列是保守的,能独立快速发展的DNA,内含子序列的研究成为动物分子系统学的基础[5]。β-纤维蛋白原第7内含子是一个富含AT的内含子,其碱基组成的变化代表着鸟类的进化[6]。β-纤维蛋白原核苷酸的多态性检测曾被用于猫科[7]等哺乳动物[8]及某些鸟类[6,9]的系统进化树分析。本文以三种鹤类为研究对象,以火烈鸟为对照,通过聚合酶链式反应以及测序实验,分析β-纤维蛋白原第七内含子的核苷酸多态性,从DNA水平来探讨三种鹤类之间的亲缘关系,寻找适合鹤科鸟类的系统分类指标。
1 材料和方法
1.1 材料来源
2~3岁龄雄性四种涉禽鸟类新鲜血液,采自哈尔滨 动 物 园 灰 鹤 (Grus grus)、 丹 顶 鹤(Grus japonensis)、 白 鹤 (Grus leucogeranus)、 火 烈 鸟(Phoenico pterus ruber)。
1.2 主要试剂
Taq Plus DNA Polymerase(购自天根生化科技(北京)有限公司);全血基因组DNA快速提取试剂盒(购自原平皓(天津)生物技术有限公司);琼脂糖凝胶DNA回收试剂盒I(离心柱型)(北京三博远志生物技术有限责任公司)18-T Vector(宝生物工程(大连)有限公司);质粒DNA小量快速制备试剂盒(杭州博日生物技术公司)等。
1.3 全血基因组DNA快速提取
取30 μL新鲜加入抗凝剂的血液,放入1.5 mL离心管。按照全血基因组DNA快速提取试剂盒说明书操作提取基因组DNA。所得DNA进行琼脂糖凝胶电泳检验,-20℃保存。
1.4 聚合酶链式反应
根据Genbank中的Grus leucogeranus betafibrinogen gene,intron 7(索引号:DQ494146) 应用primer5.0软件设计引物,其序列如表1,生工生物工程(上海)股份有限公司合成。

表1 引物序列和产物长度Table 1 Primer sequences and product length
用PCR仪扩增,94℃变性,55℃退火,72℃延伸,35个循环。PCR产物经1.2%琼脂糖凝胶电泳后,按照琼脂糖凝胶DNA纯化试剂盒操作方法纯化目的基因。
1.5 PCR产物克隆及序列测定
将PCR产物克隆入pMD18-T载体中,筛选阳性克隆,由生工生物工程(上海)股份有限公司对目的片段测序。
2 实验结果
2.1 β-纤维蛋白原第七内含子基因扩增结果
通过PCR方法扩增β-纤维蛋白原第七内含子基因,经1.2%的琼脂糖电泳。结果表明(见图1),目的条带特异、清晰,与实验设计目的片段大小一致(565 bp)。
2.2 β-纤维蛋白原第七内含子基因片段序列分析
β-纤维蛋白原第七内含子基因克隆产物测序结果为:灰鹤 (Grusgrus)565 bp、丹顶鹤(Grus japonensis)566 bp、白鹤(Grus leucogeranus)565 bp、火烈鸟(Phoenico pterus ruber)562 bp。测序结果利用DNAMAN软件进行比对(图2)。由图可知,三种鹤类β-纤维蛋白原第七内含子基因序列中第23、44、116、189位核苷酸为多态性位点。

图1 四种涉禽β-纤维蛋白原第七内含子基因PCR扩增产物Fig.1 The PCR amplification products for intron of β-fibrinogen gene of four waders
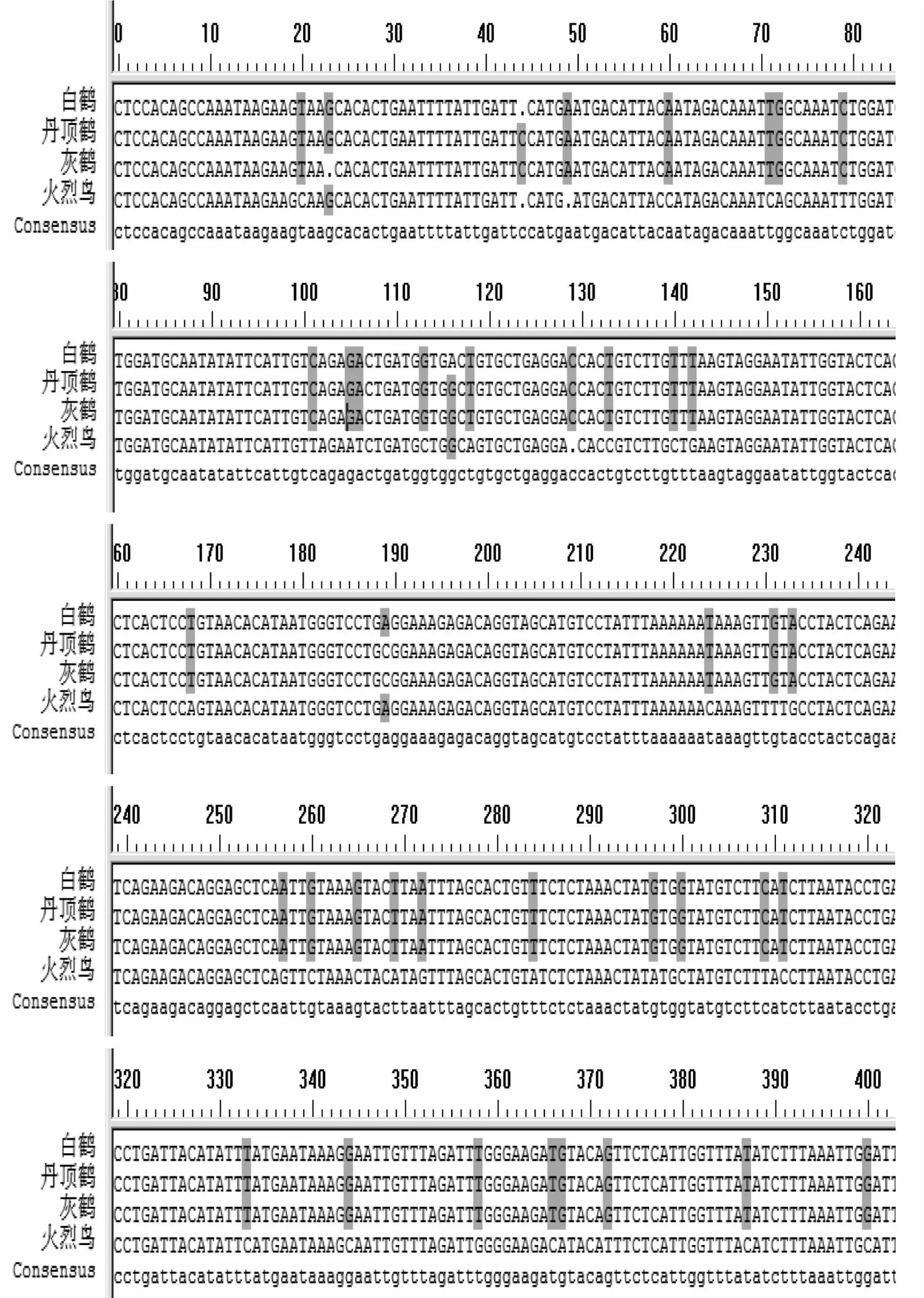
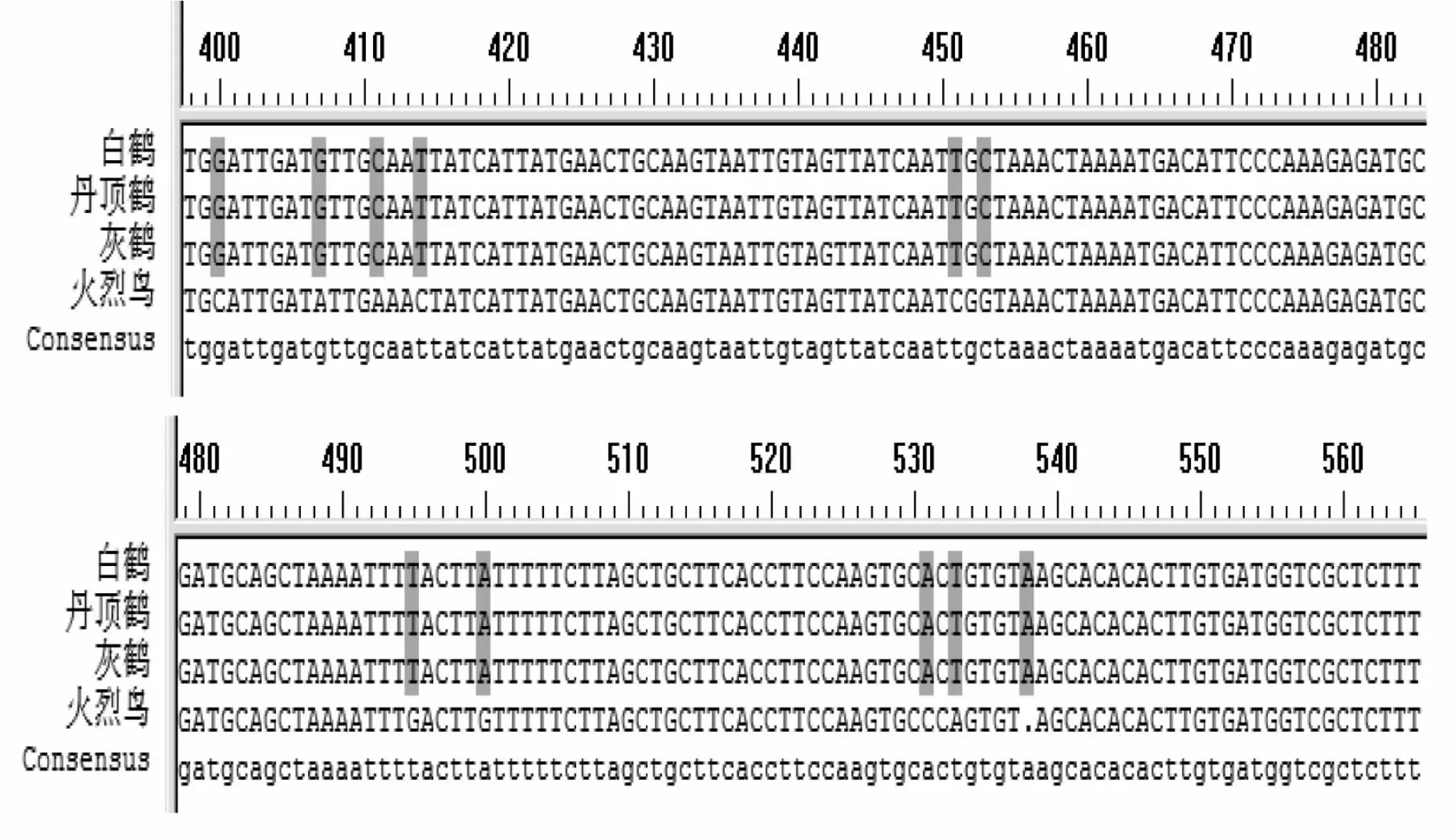
图2 四种涉禽β-纤维蛋白原第七内含子基因序列比对分析Fig.2 Comparation analysis for intron 7 of β-fibrinogen gene of four waders
2.3 β-纤维蛋白原第七内含子基因片段序列相似系数和遗传距离分析
四种涉禽β-纤维蛋白原第七内含子核苷酸序列分析结果(图3)。丹顶鹤与灰鹤核苷酸相似度最高,距离最近;火烈鸟与灰鹤核苷酸相似度最低,距离最远。

图3 四种涉禽β-纤维蛋白原第七内含子核苷酸序列相似系数和遗传距离分析Fig.3 Similarity coefficient and genetic distance analysis for intron 7 of β-fibrinogen gene of four waders
2.4 β-纤维蛋白原第七内含子基因序列聚类分析
利用DNAStar软件对三种鹤类和火烈鸟的β-纤维蛋白原第七内含子基因序列进行聚类分析,用距离法构建进化树,推演β-纤维蛋白原第七内含子的系统发生关系(图4)。丹顶鹤与灰鹤亲缘关系最近,遗传距离系数为0.08;火烈鸟与鹤类亲缘关系很远,遗传距离系数为0.972。

图4 四种涉禽β-纤维蛋白原第七内含子基因序列聚类关系Fig.4 Clustering relationship analysis for intron 7 of β-fibrinogen gene between four waders
3 讨论
本实验检测了三种鹤类和火烈鸟的β-纤维蛋白原第七内含子基因序列,并进行了序列和系统进化分析。结果表明三种鹤类β-纤维蛋白原第七内含子核苷酸序列距离在0.4以内,其中丹顶鹤和灰鹤基因序列相似性最高为99.8%,亲缘关系最近,遗传距离系数为0.08;与葛运生、陈亮等[8]2004年报道的实验结果一致。火烈鸟与鹤类亲缘关系最远,遗传距离系数为0.972。其中白鹤与火烈鸟基因序列相似性为91.2%;丹顶鹤与火烈鸟基因相似性为91.0%;灰鹤与火烈鸟基因相似性最低为90.8%。这些为鹤类的系统进化分析提供了重要实验数据。
由于β-纤维蛋白原第七内含子具有较高的属种保守性,其序列的缺失和置换在解决亲缘关系较近的分类阶元的系统关系方面具有重要价值。鹤类的β-纤维蛋白原第七内含子的核苷酸多态性,主要发生缺失和颠换(图2),这与相关的系统发育树和基因组进化分析中的结果一致[9]。与利用线粒体[10]进行亲缘关系分析一样,β-纤维蛋白原核苷酸序列的缺失和替换是构建鹤科鸟类分子进化树和鉴定亲缘关系的一个有效方法。
[1]李拥军,赵万里,丁家桐.我国鹤类的保护和研究[J].动物科学和动物医学,2000,17(1):13-15.
[2]李晓明,刘学昌.挠力河流域丹顶鹤、白枕鹤种群动态及其与环境关系研究[J].湿地科学,2005,3(2):127-131.
[3]赵春霞,杨焕民,李鹏,等.RAPD技术及其在畜禽育种中的应用[J].黑龙江八一农垦大学学报,2004,16(2):61-64.
[4]David H.Bos,David Posada.Using models of nucleotide evolution to build phylogenetic trees[J].Developmental&Comparative Immunology,2005,29(3):211-227.
[5]Thomas M.Prychitko, William S.Moore.Comparative evolution of the mitochondrial cytochrome b gene and β-fibrinogen intron 7 in woodpeckers[J].Molecular Biology and Evolution,2000,17(7):1101-1111.
[6]Thomas M.Prychitko,William S.Moore.Alignment and phylogenetic analysis of β-fibrinogen intron 7 sequences among avian orders reveal conserved regions within the Intron[J].Molecular Biology and Evolution,2003,20(5):762-771.
[7]Li Yua,Yaping Zhang.Phylogenetic studies of pantherine cats (Felidae) based on multiple genes,with novel application of nuclear β-fibrinogen intron 7 to carnivores[J].Molecular Phylogenetics and Evolution,2005,35(2):483-495.
[8]Conrad A.Matthee,Geeta Eick,Sandi Willows-Munro,et al.Indel evolution of mammalian introns and the utility of non-coding nuclear markers in eutherian phylogenetics[J].Molecular phylogenetics and evolution,2007,42(3):827-837.
[9]Lukasz Pasko,Per G.P.Ericson,Andrzej Elzanowski.Phylogenetic utility and evolution of indels:A study in neognathous birds [J]. Molecular Phylogenet and Evolution,2011,61(3):760-771.
[10]葛云生,陈亮,沈明山,等.几种鹤类线粒体DNA的差异和亲缘关系分析[J].厦门大学学报:自然科学版,2004,43(5):718-723.
[11]李庆伟,田春宇,李爽.鹰科四种鸟类线粒体DNA的差异和分子进化关系研究[J].遗传,2001,23(6):529-534.
[12]Morgan-Richards M,Trewick SA,Bartosch-Harlid A,et al.Bird evolution:testing the Metaves clade with six new mitochondrial genomes [J].BMC evolutionary biology,2008,23(8):20.
猜你喜欢
作物学报(2022年2期)2022-11-06 12:11:02
中国现代中药(2021年9期)2021-11-16 07:43:32
内蒙古师范大学学报(自然科学汉文版)(2021年3期)2021-06-01 08:10:30
生物工程学报(2019年6期)2019-07-10 08:38:38
生物学通报(2019年1期)2019-02-15 16:33:43
天然产物研究与开发(2018年10期)2018-11-06 07:43:46
生物学通报(2018年12期)2018-10-10 06:52:36
小天使·六年级语数英综合(2018年5期)2018-10-10 00:00:00
广东农业科学(2017年5期)2017-08-29 10:37:26
小天使·六年级语数英综合(2016年6期)2016-05-14 21:40:55
