
TRAIL诱导恶性胶质瘤LN215细胞凋亡抵抗的机制
2013-05-29 09:17:44张艳春于洪泉赵东海刘兴吉
中国老年学杂志 2013年6期
张艳春 于洪泉 王 猛 赵东海 金 宏 齐 玲 刘兴吉
(吉化集团公司总医院麻醉科,吉林 吉林 132013)
恶性胶质瘤目前尚缺乏理想的治疗方法〔1〕。肿瘤坏死因子相关凋亡诱导配体(TRAIL)作为一种新型的抗肿瘤药物,在美国已进入临床Ⅱ期实验阶段〔2〕。但大部分恶性胶质瘤细胞株对TRAIL诱导凋亡发生抵抗〔3〕,其机制仍然不清。本文对恶性胶质瘤细胞LN215细胞和单克隆细胞进行研究,为TRAIL作为肿瘤治疗新药提供实验依据。
1 材料与方法
1.1 主要试剂 重组人TRAIL(rhTRAIL)(Peprotech公司);DMEM/F12培养基、小牛血清(Gibco公司);对硝基苯(上海生物工程技术公司);兔抗人第二个线粒体激活因子(SMAC)多克隆抗体(Biomol公司);兔抗人BH3结构域凋亡诱导蛋白(BID)多克隆抗体(Invitrogen公司);兔抗人肌动蛋白(actin)多克隆抗体(Sigma公司);过氧化物酶标记IgG第二抗体(Jackson Immunoresearch公司)。
1.2 细胞培养及分离单克隆细胞 将LN215细胞接种于含10%小牛血清的DMEM/F12培养基中培养,待细胞汇合至80%~90%,用0.25%胰酶消化,台盼蓝计数后稀释。按1~2个/孔细胞密度接种于96孔板中进行培养。培养条件为5%CO2,37℃孵箱。每天观察生长情况,标记出含单个细胞的孔。待其汇合,转入24孔板中进行培养。每周换液2次,90%汇合后转入65 mm培养皿中进行培养。
1.3 酸性磷酸酶法检测细胞凋亡的百分率 待细胞融合至85%,0.25%胰酶消化,台盼蓝计数活细胞后,以1×104个/孔细胞密度接种于96孔板中,培养24 h,将细胞分为11组,加入TRAIL(0.01,0.03,0.1,0.3,1,3,10,30,100,300 ng/ml),对照组使用基础培养基。24 h后小心弃上清,磷酸盐缓冲液(PBS)洗去培养基,每孔加入酸性磷酸酶底物检测液(对硝基苯新鲜配制)100μl,继续培养90 min,加入终止液,室温下孵育5 min。酶标仪405 nm处测定各孔光吸收值。按照公式计算细胞凋亡率=(1-药物组光吸收值/对照组光吸收值)×100%。
1.4 Western印迹法检测蛋白的表达 收集细胞,3 000 r/min离心后弃上清。PBS洗涤细胞离心后弃上清,每个管中加入50μl细胞裂解液,12 000 r/min 4℃ 离心 15 min,弃去沉淀,Braford测定法检测样品蛋白浓度,十二烷基硫酸钠-聚丙烯酰胺凝胶(SDS-PAGE)电泳分离样品,4℃转膜封闭,加入一抗4℃过夜。加入二抗,室温、避光孵育1 h,胶片曝光拍照。
1.5 统计学分析 采用SPSS11.0软件进行分析,计量资料以±s表示,行t检验。
2 结果
2.1 LN215细胞及单克隆细胞的培养 恶性胶质瘤LN215细胞经复苏后培养,细胞呈现长梭形,有的可见明显的突起。观察96孔板内含单细胞孔细胞的生长状态发现:虽然细胞来自于同一株细胞,但各单克隆细胞生长速度不同、形态也不全相同。待细胞生长至90%时,挑选出生长速度与形态均不相同的32株细胞,用胰酶消化转入24孔板生长,细胞生长状态良好。比较后发现有5株单克隆细胞生长形态、生长速度不同,编号分别是1、6、17、13和32。这5株细胞用于后续实验。
2.2 LN215细胞和单克隆细胞对TRAIL诱导凋亡的敏感性LN215和5株单克隆细胞经不同浓度TRAIL作用后,细胞凋亡率在 -10.2% ~55.7%之间波动,LN215、1、6、17、13 和 32 细胞的最大凋亡率分别为12.1%、3.48%、17.4%、4.85%、55.7%和28.1%,其中6、13和32经TRAIL作用后发生凋亡较LN215更明显,而1和7则相反,以13对TRAIL诱导凋亡最为敏感。见图1。
2.3 LN215细胞和单克隆细胞TRAIL相关蛋白的表达TRAIL相关蛋白均有表达。LN215细胞和单克隆细胞表达DR4与细胞凋亡百分率不相关;BID、SMAC表达与细胞凋亡百分率明显相关,6、13、32较1和17表达量明显增加,尤其是13最为明显。见图2。
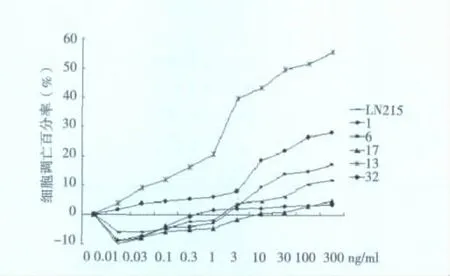
图1 TRAIL诱导LN215及单克隆细胞发生凋亡情况
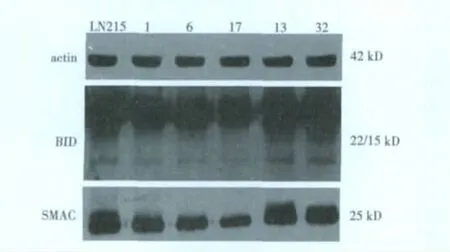
图2 恶性胶质瘤LN214细胞及单克隆细胞TRAIL相关蛋白的表达
3 讨论
胶质瘤是起源于中枢神经系统的最常见肿瘤,由于其发病隐匿,一经发现病理诊断就是Ⅲ~Ⅳ期胶质瘤,给临床治疗带来了很大困难。并且由于脑胶质瘤在脑内呈弥散分布〔4〕,手术难以清除。这可能是由于激活细胞生长途径和/或抑制细胞凋亡途径而对治疗产生抵抗〔5〕。TRAIL可以诱导胶质瘤细胞凋亡〔6,7〕,TRAIL诱导凋亡的非线粒体途径需要线粒体途径的补充,由线粒体途径通过(caspase 8)间接激活BID,再依次与Bax和Bak作用,改变了线粒体膜的生物学特性,促使细胞色素C和SMAC等线粒体蛋白释放。在胞质内,SMAC与相关蛋白作用解除抑制因子的抑制作用,使TRAIL诱导的非线粒体途径得以进行〔8,9〕,细胞发生凋亡。
根据肿瘤发生可能来源于多个细胞的突变,本实验利用细胞梯度稀释方法由LN215分离出5株单克隆细胞。LN215细胞和单克隆细胞BID、SMAC表达与细胞凋亡百分率明显相关。因此,笔者认为LN215和单克隆细胞经TRAIL诱导凋亡百分率与BID和SMAC表达量密切相关,说明TRAIL诱导LN215和单克隆细胞可能是通过BID和SMAC引起,表达量的减少可能会引起TRAIL诱导凋亡抵抗,但更进一步的机制仍待研究。
1 Stupp R,Mason WP,van den Bent MJ,et al.Radiotherapy plus concomitant and adjuvant temozolomide for glioblastoma〔J〕.N Engl J Med,2005;352(10):987-96.
2 Bellail AC,Qi L,Mulligan P,et al.TRAIL agonists on clinical trials for cancer therapy:the promises and the challenges〔J〕.Revi Rec Clin Trials,2009;(4):34-41.
3 Li YC,Tzeng CC,Song JH,et al.Genomic alterations in human malignant glioma cells associate with the cell resistance to the combination treatment with tumor necrosis factor-related apoptosis-inducing ligand and chemotherapy〔J〕.Clin Cancer Res,2006;12(9):2716-29.
4 Holland EC.Glioblastoma multiforme:the terminator〔J〕.Proc Natl Acad Sci USA,2000;97(12):6242-44.
5 Lefranc F,Brotchi J,Kiss R.Possible future issues in the treatment of glioblastomas:special emphasis on cell migration and the resistance of migrating glioblastoma cells to apoptosis〔J〕.J Clin Oncol,2005;23(10):2411-22.
6 南栗岩,齐 玲,肖振晶,等.原代培养的恶性胶质瘤细胞表达死亡受体与TRAIL抵抗的关系〔J〕.中国老年学杂志,2011;18(1):3591-2.
7 齐 玲,于洪泉,丁丽娟,等.Caspase-8在胶质母细胞瘤抵抗TRAIL诱导凋亡中的作用〔J〕.吉林大学学报(医学版),2011;37(4):612-6.
8 Li H,Zhu H,Xu CJ,et al.Cleavage of BID by caspase8 mediates the mitochondrial damage in the Fas pathway of apoptosis〔J〕.Cell,1998;94(8):491-501.
9 Du C,Fang M,Li Y,et al.Smac,a mitochondrial protein that promotes cytochrome c-dependent caspase activation by eliminating IAP inhibition〔J〕.Cell,2000;102(1):33-42.
猜你喜欢
中学生理科应试(2024年6期)2024-01-01 00:00:00
核科学与工程(2022年3期)2022-10-18 01:25:14
中国现代医药杂志(2020年10期)2020-12-14 07:20:14
广州化工(2020年6期)2020-04-18 03:30:20
现代矿业(2018年9期)2018-10-16 09:37:02
现代检验医学杂志(2016年3期)2016-11-15 01:59:28
医学研究杂志(2015年3期)2015-06-10 06:41:52
特产研究(2015年1期)2015-04-12 06:36:20
西华师范大学学报(自然科学版)(2015年3期)2015-02-27 15:31:22
天然气与石油(2014年2期)2014-07-16 11:01:18
