
光叶紫花苕种子贮藏蛋白分析
2013-05-09 11:13王清郦孙启忠敖学成
草原与草坪 2013年4期
柳 茜,王清郦,傅 平,孙启忠,敖学成
(1.四川省凉山州畜科所,四川 西昌 615042;2.中国农业科学院研究生院,北京 100081;3.中国农业科学院草原研究所,内蒙古 呼和浩特 010010)
光叶紫花苕(Vicia villosavar.glabresens)为一年生草本植物,原产于美国俄勒冈州,20世纪40年代自美国引进南京,现广泛种植于安徽、山东、河南、湖北、云南、四川、贵州等省[1]。光叶紫花苕适应性广,耐寒,耐旱,在田间、果园、山坡、生荒地上均可种植[2]。光叶紫花苕具有较高的饲用价值,粗蛋白含量在20%以上,氨基酸、矿物质、维生素含量比较丰富,是牛、猪、鸡的优质饲料[3-7]。此外,光叶紫花苕还是良好的绿肥和覆盖作物,能改良土壤肥力,提高土壤固氮能力以及后续作物的产量和品质[8,9],同时也是良好的蜜源植物[2]。凉山光叶紫花苕(Vicia villosavar.glabresenscv.Liangshan)是凉山州从云南引进的光叶紫花苕在长期推广中选育出的地方品种,适应性强,在海拔2 500~3 200m的地区都能种植[3],是高寒山区冬春季节的优质牧草。
种子贮藏蛋白是植物体中一类表达率高、能在种子内大量积累且不易降解的蛋白质[10]。贮藏蛋白可分为醇溶蛋白、谷蛋白、清蛋白和球蛋白等,这些蛋白质的组成由遗传决定,不受环境影响,其组分上的差异可以反映出基因型的不同,因而被称为品种的生化“指纹”[11]。研究种子贮藏蛋白的方法有多种,其中蛋白质电泳谱带具有较高的多态性、专一性和稳定性,在植物的遗传生化标记、品种鉴定、种群(居群)间的遗传变异以及遗传结构的研究等方面得到广泛应用[12,13]。试验采用十二烷基磺酸钠聚丙烯酰胺凝胶电泳(SDSPAGE)技术对凉山州境内5个居群的光叶紫花苕的种子贮藏蛋白进行研究,从生化水平上探讨凉山光叶紫花苕的遗传多样性,为凉山州种质资源的研究和开发利用提供技术参考依据。
1 材料和方法
1.1 材料
试验材料采自凉山州境内的西昌、盐源、普格、普格小兴场和德昌5个地区光叶紫花苕的自然居群。每个居群随机选取5个单株混合作为该居群的样品。各居群的生态因子情况见表1。
1.2 试验方法
参照文献[14]SDS-PAGE垂直平板电泳方法,分别对各居群种子贮藏蛋白(清蛋白、谷蛋白、醇溶蛋白和盐溶蛋白)及总贮藏蛋白进行电泳,并分析其多态性。

表1 光叶紫花苕居群的样品来源Table 1 The Source of Vicia villosa Roth var.glabrescens cv.Liangshan samples
1.2.1 种子贮藏蛋白的提取 取10粒种子洗净擦干,放入研钵中研碎,置于1.5mL离心管中,加入蛋白提取液[15-18]。
①清蛋白提取液:去离子水500μL。
②盐溶蛋白提取液:加200μL脱脂剂(氯仿∶甲醇∶丙酮=2∶1∶1),振荡混匀,脱脂2h(室温),10 000 r/min离心10min,弃上清液后风干,加入500μL盐溶蛋白提取液(0.25mmol/L Na3PO4、2mol/L NaCl,pH =7.0)。
③醇溶蛋白提取液:75%乙醇溶液500μL。
④谷蛋白提取液:0.2%NaOH溶液500μL。
⑤贮藏蛋白提取液:5mL 10%SDS,1.5mL 2-ME,1.25g溴酚兰,3.44mL 甘油,2.0mL 1mol/L Tris-Hcl pH 6.8,加双蒸水定容到25mL;500μL。
室温振荡浸提2h,10 000r/min离心10min,取上清液置于4℃冰箱备用。以1∶1的比例加入上样缓冲液,煮沸5min后点样。
1.2.2 SDS-PAGE电泳 SDS-PAGE的浓缩胶浓度3%,分离胶浓度12%。各类贮藏蛋白及总的贮藏蛋白的上样量10μL,标准蛋白D532S(分子量6.5~200 kDa),上样量5μL,用北京六一电泳仪器厂DYZ-28B型电泳槽,浓缩时电压恒压200V,分离时电流恒流60 mA电泳,室温下电泳7~8h至指示剂跑到距底部1cm处,电泳结束后剥下胶片,染色3~10h,脱色至条带清晰为止,照相并进行统计[18]。
1.3 统计分析
SDS-PAGE中蛋白单体分子量的电泳迁移率主要取决于它的分子量,而与所带电荷和形状无关,单体电泳迁移率与其蛋白单体分子量的关系为lgMW=lgK-bm=K1-bm。其中MW为蛋白单体分子量,m为蛋白单体迁移率,K,K1和b为常数。相对迁移率(Rf)=脱色后蛋白质移动距离×染色前分离胶长/染色前指示剂移动距离×脱色后分离胶长。根据已知分子量的标准蛋白(Marker)在相同电流条件下的迁移率,作出分子量和迁移率的标准曲线,然后计算出待测蛋白单体的分子量。
根据电泳结果对清晰蛋白质条带的有无进行标记,有为1,无为0,得出(0,1)矩阵。利用 Popgene 1.31软件进行Nei’s遗传距离(D)和遗传一致度(I)分析。利用NTSYS-pc 2.1系统进行聚类分析[19]。
2 结果与分析
2.1 贮藏蛋白谱带遗传多样性分析
利用SDS-PAGE对5个不同居群的光叶紫花苕种子贮藏蛋白进行电泳,获得清晰稳定的遗传蛋白图谱(图1)。
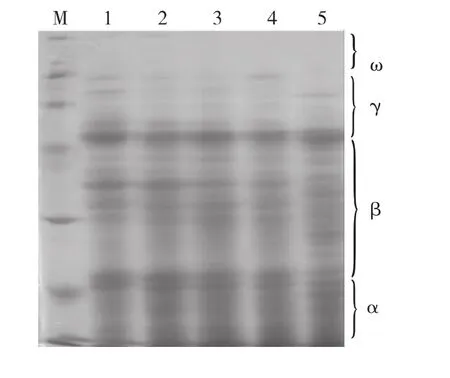
图1 5个不同居群光叶紫花苕的贮藏蛋白电泳图谱Fig.1 Electrophoregrams of seed storage proteins of 5 populations of Vicia villosa var.glabrescens cv.Liangshan
5个居群之间的贮藏蛋白谱带存在差异,共检测到31条蛋白谱带,其中,11条谱带为共有带,包括第3号、7号、8号、14 号、16 号、22 号 、25 号 、26 号、28 号、30号、31号;其余20条为不同程度表达的多态性谱带,多态性数目占总条带的65.52%。西昌居群有19条带,盐源居群有22条带,普格小兴场居群有20条带,普格居群有17条带,德昌居群有22条带。普格小兴场居群有1条特有带,德昌居群有8条特有带。谱带分子量在11.09~100.09kDa,相对迁移率在0.16~0.84。根据谱带相对迁移率由小到大(分子量由大到小),将种子贮藏蛋白图谱分为α、β、γ、ω区域。其中α区相对迁移率为0.7~1.0,β区相对迁移率为0.3~0.7,γ区相对迁移率为0.1~0.3,ω区相对迁移率为0~0.1。贮藏蛋白只在α、β、γ区域有条带出现,其中α区有6条,β区有19条,γ区有6条,ω区无谱带出现,表明光叶紫花苕种子贮藏蛋白主要集中在β区。
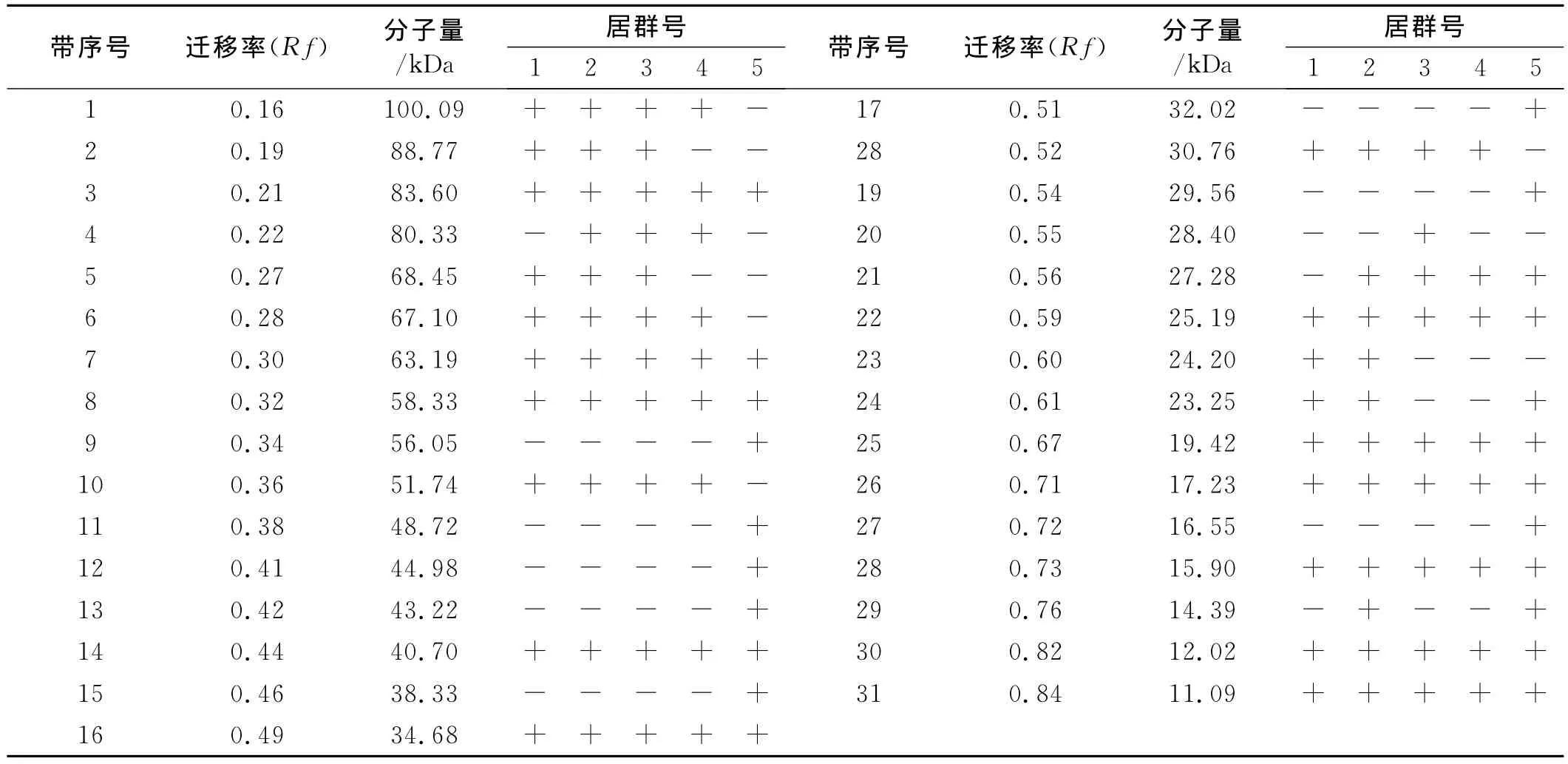
表2 5个不同居群光叶紫花苕贮藏蛋白电泳各谱带的迁移率和蛋白分子量Table 2 Frequency of bands and molecular weight of seed storage proteins of 5populations of Vicia villosa var.glabrescens cv.Liangshan
光叶紫花苕5个居群贮藏蛋白的Nei’s的遗传距离和遗传一致度见表3,西昌与盐源居群、普格小兴场与普格居群的遗传距离最小,均为0.101 8,普格小兴场与德昌居群遗传距离最大,为0.869 0;普格小兴场与德昌居群遗传一致度最小,为0.419 4,西昌与盐源居群、普格小兴场与普格居群的遗传一致度最大,为0.903 2。
5个居群的贮藏蛋白谱带遗传相似系数在0.13~0.90,平均达0.60,遗传相似性较高,表明遗传多样性较低,遗传基础较窄。居群1与2、3与4谱带相似系数最高,达到0.90,说明居群间亲缘关系较近;居群5与1、2、3、4号居群的遗传相似系数小,分别为0.21、0.27、0.13、0.34,表明居群间亲缘关系较远。从聚类分析结果可以看出(图2),在遗传相似系数为0.47时5个居群分为2大类,第1类为1、2、3、4号居群,5号居群单独聚为1类。
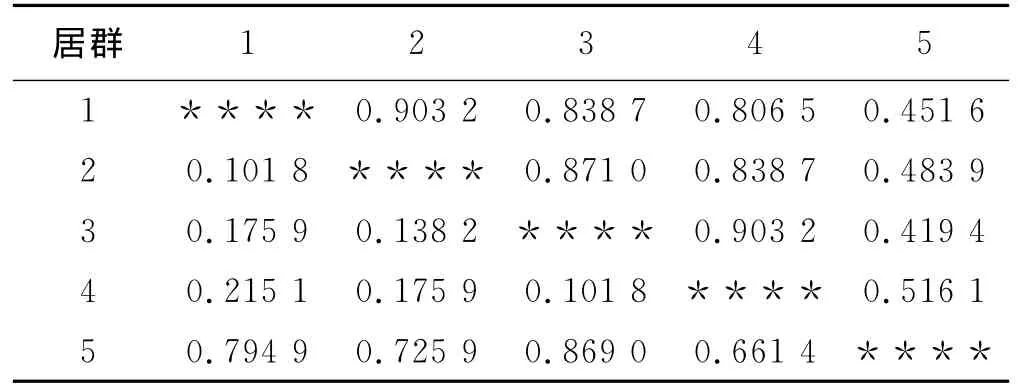
表3 贮藏蛋白Nei’s遗传一致度(右上角)和遗传距离(左下角)Table 3 Nei's of genetic identity and genetic distance of seed storage proteins
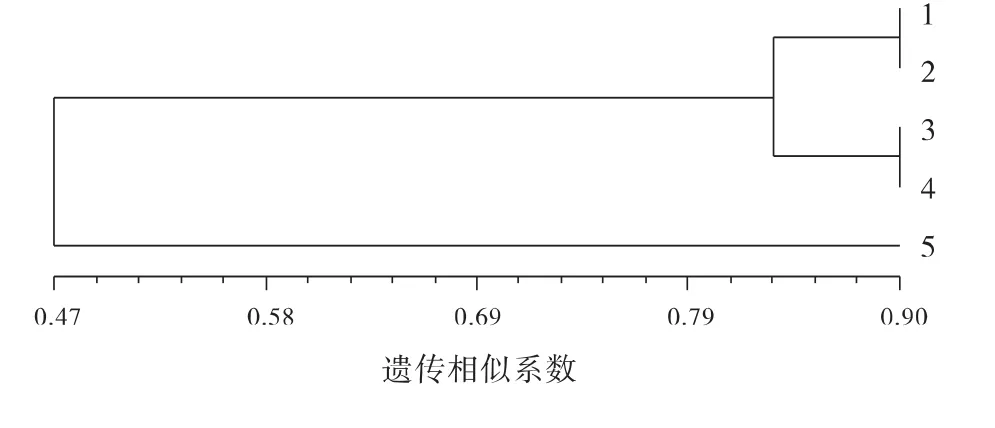
图2 5个居群光叶紫花苕的贮藏蛋白聚类图Fig.2 Dendrogram based on Nei’s genetic distance of 5populations of Vicia villosa var.glabrescens cv.Liangshan
2.2 盐溶蛋白谱带遗传多样性分析
5份供试材料的盐溶蛋白谱带存在差异,共检测出22条蛋白谱带,其中13条谱带为稳定表达谱带,表达率为100%;其余9条为不同程度的多态性谱带,多态性数目占总条带的40.92%(图3)。5(德昌居群)有8号、14号两条特有谱带,1(西昌居群)有17号1条特有谱带。其中α区有5条,β区有12条、γ区有5条,ω区无谱带出现,分子量大小13.50~95.04kDa(表4)。
5个不同居群光叶紫花苕盐溶蛋白的Nei’s的遗传距离和遗传一致度见表5,2(盐源居群)与4(普格居群)的遗传距离为0最小,5(德昌居群)与2(盐源居群)、4(普格居群)的遗传距离最大,为0.452 0;5(德昌居群)与2(盐源居群)、4(普格居群)的遗传一致度最小(0.636 4),4(普格居群)与2(盐源居群)的遗传一致度最大(1.000 0)。

图3 5个不同居群光叶紫花苕的盐溶蛋白电泳图谱Fig.3 Electrophoregrams of salt-soluble proteins of 5populations of Vicia villosa var.glabrescens cv.Liangshan
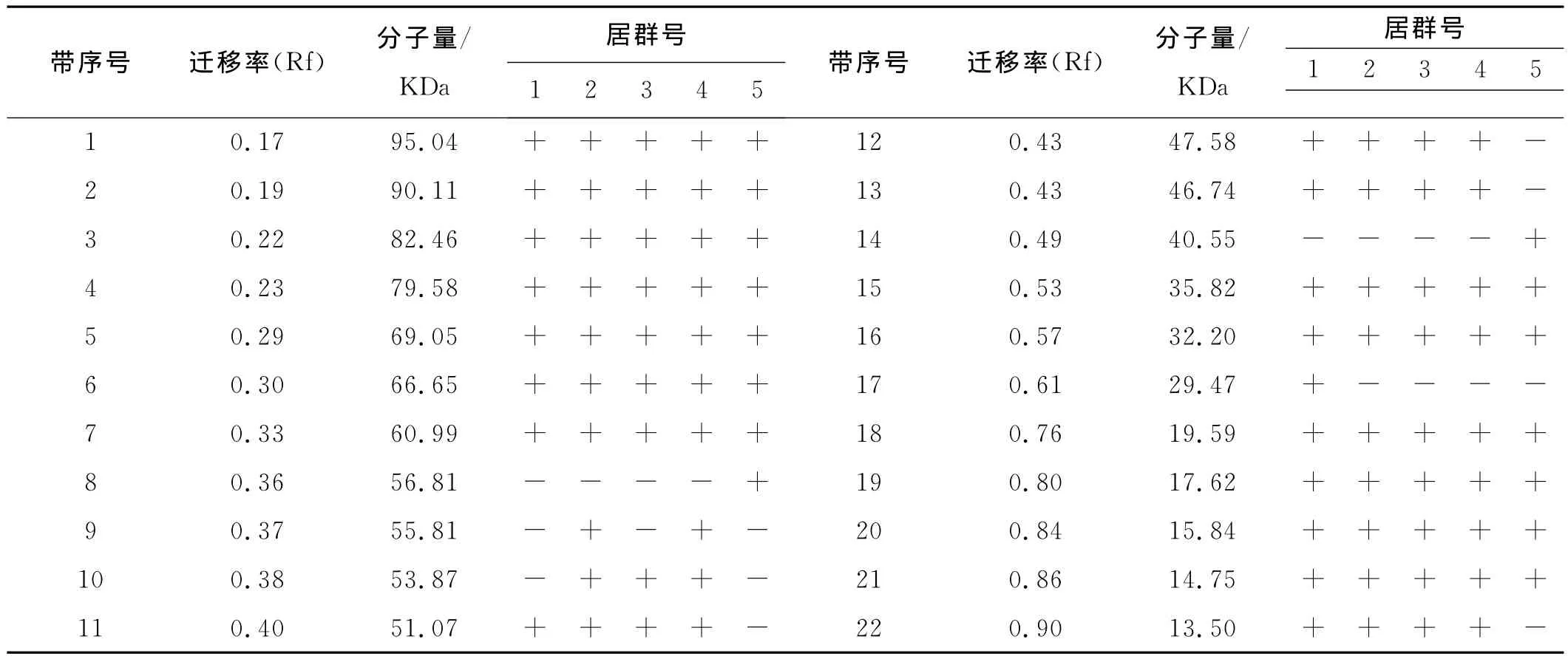
表4 5个不同居群光叶紫花苕盐溶蛋白电泳各谱带的迁移率(Rf)和蛋白分子量Table 4 Frequency of bands and molecular weight of salt-soluble proteins of 5populations of Vicia villosa var.glabrescens cv.Liangshan
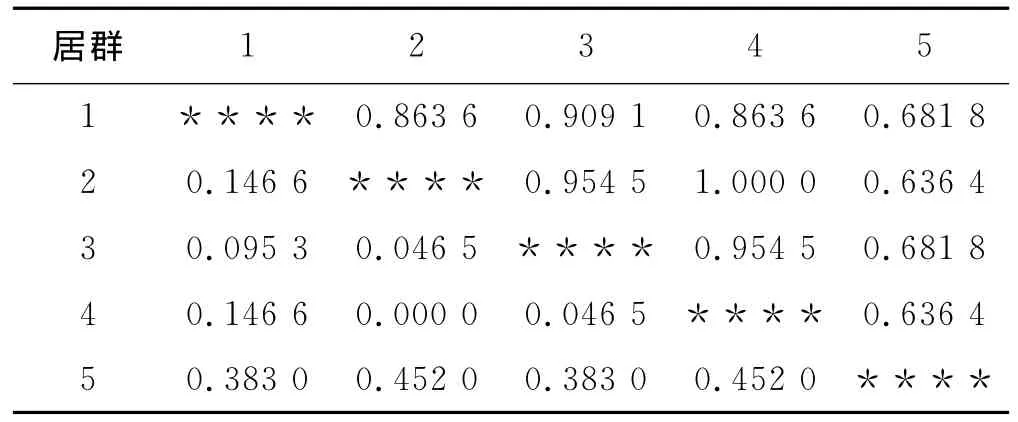
表5 盐溶蛋白Nei’s遗传一致度(右上角)和遗传距离(左下角)Table 5 Nei's of genetic identity and genetic distance of salt-soluble proteins
5个居群的盐溶蛋白谱带遗传相似系数在0.55~1.00,平均达0.78,谱带相似系数最高的是居群2与4,达到1.00,居群间亲缘关系最近;居群5与2、4号居群的遗传相似系数较低,为0.55,居群间亲缘关系最远。从聚类分析结果可以看出(图4),在遗传相似系数为0.66时5个居群分为2大类,第1大类又可分2个亚类,2、3、4号居群聚为1类,1号居群单独为1类。5号居群单独聚为第2大类。
2.3 清蛋白、谷蛋白和醇溶蛋白电泳结果
经电泳染色脱色后,点样孔加入醇蛋白的泳道无清晰谱带出现,点样孔加入清蛋白、谷蛋白的泳道中出现几条痕迹很浅的条带,由于蛋白含量过少故对此不做过多讨论。
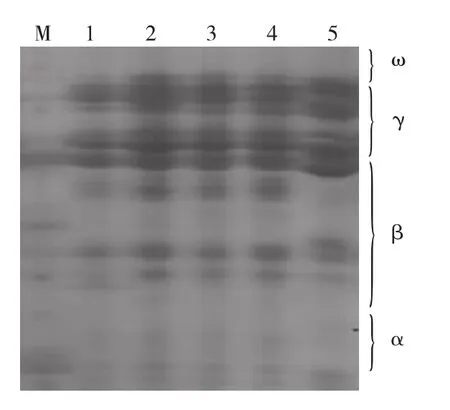
图4 5个不同居群光叶紫花苕的盐溶蛋白聚类图Fig.4 Dendrogram based on Nei’s genetic distance of 5 Vicia villosa var.glabresens cv.Liangshan
3 讨论与结论
SDS-PAGE凝胶电泳的电泳速度与蛋白分子量大小有关而与电荷和形状无关。因其具有电泳结果稳定、不受贮藏年限和外界环境影响等优点而得到广泛应用[20]。李拥军等[20]利用SDS-PAGE方法对我国18个苜蓿地方品种和北美9个苜蓿基本种质来源的代表品种及14份豆科牧草种(品种)种子贮藏蛋白进行了分析,结果表明贮藏蛋白谱带差异较小;闫伟红等[21]对引种栽培筛选出的5种12个冰草属植物居群醇溶蛋白进行遗传结构分析,发现同一物种不同来源材料间遗传差异较小。
对5个不同居群光叶紫花苕的蛋白电泳谱带进行统计,不同提取液所获得的蛋白谱带各不相同。其中使用75%乙醇溶液提取的种子醇溶蛋白含量甚微或无,使用去离子水和0.2%NaOH溶液提取的种子清蛋白和谷蛋白只出现几条痕迹很浅的条带。利用盐溶蛋白和贮藏蛋白提取液所得的总蛋白谱带的分子量介于11.09~100.09kDa,相对迁移率在0.17~0.90。盐溶蛋白的蛋白谱带总数为22条,多态率为40.92%;贮藏蛋白的为31条,多态率为65.52%,说明5个居群的盐溶蛋白的条带位点少于贮藏蛋白,种内变异小于贮藏蛋白。聚类结果表明,利用盐溶蛋白和贮藏蛋白获得的蛋白质图谱能区分鉴别5个不同居群的光叶紫花苕。供试材料分为2类:1)西昌、盐源、普格小兴场、普格居群;2)德昌居群。表明同一物种不同生态居群的遗传差异较小,与闫伟红研究结果相似[21]。凉山光叶紫花苕具有较高的遗传保守性,在不同生态区都能保持较稳定的遗传性,这可能与光叶紫花苕为自花授粉植物有关。德昌居群种子千粒质量比其他居群小,但植物学特性表现一致,是否是光叶紫花苕新类型,还需要结合其他手段做进一步的研究证明。
[1]中国农业百科全书总编辑委员会农作物卷编辑委员会.中国农业百科全书·农作物卷上[M].北京:农业出版社,1991:214.
[2]中国农业百科全书总编辑委员会养蜂卷编辑委员会.中国农业百科全书养蜂卷[M].北京:农业出版社,1993:112.
[3]全国牧草品种审定委员会.中国牧草登记品种集[M].北京:中国农业大学出版社,1999:181.
[4]赵庭辉,李树清,邓秀才.高海拔地区光叶紫花苕不同生育时期的营养动态及适宜利用期[J].中国草食动物,2010,30(3):4-6.
[5]邹文能,董建强,朱辉鸿,等.试论光叶紫花苕在高寒贫困山区的推广应用[J].甘肃农业大学学报,2009,44(1):13-15.
[6]杨玉能,夏先林,田茂林.用紫花苕代替育肥肉牛精料中豆粕试验[J].草业科学,2010,10(24):56-71.
[7]王德凤,徐国录,周鹏,等.光叶紫花苕对杂交肉牛生长代谢的影响研究[J].贵州畜牧兽医,2010,34(5):7-9.
[8]侣国涵,吴文昊,梅东海,等.不同光叶紫花苕子翻压量对烤烟产量和品质的影响[J].中国烟草科学,2011,10(32):82-86.
[9]卯升华,董恩省,彭瑶.光叶紫花苕对土壤肥力及后续作物产量的影响[J].现代农业科技,2007(11):102-104.
[10]胡志昂,王洪新.蛋白质多样性和品种鉴定[J].植物学报,1991,33(7):556-2564.
[11]兰海燕,李立会.蛋白质凝胶电泳技术在作物品种鉴定中的应用[J].中国农业科学,2002,35(8):916-920.
[12]夏兰琴,蒋尤泉,阎福林.扁蓿豆遗传多样性的研究[J].中国草地,1997(2):30-35.
[13]郭江波,赵来喜.中国苜蓿育成品种遗传多样性及亲缘关系研究[J].中国草地,2004,26(1):9-14.
[14]汪家政,范明.蛋白质技术手册[M].北京:科学出版社,2008.
[15]赵杨,骈瑞琪,陈晓阳,等.二色胡枝子种子储藏蛋白多样性研究[J].西北植物学报,2007,27(9):1767-1771.
[16]孙雁,朱有勇,朱永平,等.蛋白质电泳在豌豆品种鉴定中的应用[J].种子,2004,23(2):24-26.
[17]毛小涛,王照兰,杜建材,等.不同品系扁蓿豆种子蛋白指纹图谱研究[J].中国草地学报,2009,31(6):45-51.
[18]闫伟红,徐柱,师文贵,等.胡枝子属植物40个野生居群种子蛋白谱带多样性研究[J].草业科学,2007,24(5):58-63.
[19]朱长志,赖德强,吴斌,等.利用种子蛋白SDS-PAGE技术进行抗热萝卜种质鉴定[J].中国蔬菜,2010(8):21-25.
[20]李拥军,苏加楷.中国苜蓿地方品种亲缘关系的研究[J].草业学报,1999,8(1):31-41.
[21]闫伟红,徐柱,李临杭,等.冰草属植物醇溶蛋白遗传分析与评价[J].草原与草坪,2010,30(1):1-6.
猜你喜欢
河北科技师范学院学报(2022年2期)2022-08-26
浙江中医药大学学报(2021年6期)2021-07-12
中国粮油学报(2020年12期)2021-01-09
中国奶牛(2020年11期)2020-12-23
扬子江诗刊(2020年3期)2020-11-17
农民致富之友(2020年16期)2020-06-19
扬子江(2020年3期)2020-06-08
草地学报(2018年5期)2018-11-07
现代园艺(2018年1期)2018-03-15
周末·校园文学(2017年17期)2017-07-08
