
不同倍性烟草杂交获得六倍体烟草植株
2013-03-20 09:16魏烨昕党江波刘超吴天姣汪卫星郭启高梁国鲁
中国烟草学报 2013年6期
魏烨昕,党江波,刘超,吴天姣,汪卫星,郭启高,梁国鲁
西南大学园艺园林学院,重庆 400716
烟草(Nicotiana tabacum)为茄科(Solanaceae)烟草属植物,在我国乃至全世界范围内广泛种植。烟草抗病育种是近年来烟草育种的重点和难点。众多烤烟品种如云烟87虽栽培广泛,但受黑胫病危害严重[1],部分烟草品种如L-8对黑胫病具较强抗性,是烤烟抗病育种的良好抗源[2]。利用抗病品种与栽培品种杂交可获得大量抗病育种资源,但是四倍体间杂交需经多次轮回杂交才能获得一定数量抗病的优良株系。在咖啡育种的研究中,经染色体数量不对称杂交,可减少抗源基因的引入,其BC1代与栽培种相似度更高[3]。可见,染色体数量不对称杂交是抗病育种中减少轮回杂交次数,加快育种进程的较好方法。因此,通过烤烟八倍体与抗黑胫病烟草品种四倍体杂交创制烟草六倍体对烟草抗黑胫病育种具积极意义。对六倍体烟草的育性进行研究也将利于对六倍体烟草的在育种中的利用。
烟草染色体数目为2n=4x=48,为异源四倍体(SSTT)。由于其为经济作物,其育种历史较长,常进行种内、种间杂交,因此诸多育种材料倍性复杂。有单体[4-6]、缺体[7-8]、双单倍体[9-11]、附加系和代换系[12-16]及其它非整倍体[17-18]等类型报道,也有部分关于烟草与其同属其它物种杂交加倍获得异源三倍体、六倍体[19]、八倍体[20]等育种中间材料的报道。但有关烟草种内六倍体(2n=6x=72)的研究尚未见报道。
本研究通过云烟87的八倍体(2n=8x=96)植株与对黑胫病具较强抗性的烟草品种L-8的四倍体(2n=4x=48)植株杂交,获得烟草六倍体(2n=6x=72),并移栽田间种植。通过观察统计该烟草六倍体花器官特征、花粉形态,测定其花粉活力,以及田间杂交试验对该烟草六倍体的育性进行初步研究,以初步了解倍性变化对烟草育性的影响及便于在育种实践中对其进行利用。
1 材料与方法
1.1 材料
L-8四倍体植株(2n=4x=48,以下称L-8)、云烟87四倍体植株(2n=4x=48,以下称云烟87)、本实验室创制的云烟87八倍体植株(2n=8x=96,以下称八倍体),云烟87四倍体×L-8四倍体F1代四倍体植株(2n=4x=48)(以下称四倍体杂种),大田种植。
1.2 方法
1.2.1 烟草六倍体的创制
于盛花期取L-8花药,干燥阴凉处使花药自然开裂,收集花粉,于4℃干燥保存。于晴朗无风的上午将L-8花粉授于八倍体的柱头上,约30 d后采集蒴果、收集种子。将收集的种子育室温保湿催芽后播种,待幼苗长至5-6叶期时取幼嫩叶片,采用改良去壁低渗火焰干燥法进行染色体制片,镜检,对幼苗的染色体数目进行鉴定。
将获得的六倍体材料栽田间以进行后续实验。
1.2.2 烟草六倍体开花状态及花粉形态观察
形态观察及测定:在盛花期,以四倍体杂种为对照,观察并测量六倍体花器官相关特征。取即将开放、花药尚未裂开、花丝自然伸长的单个花序,测量花长度,中部直径,子房纵、横径,花柱长度、直径,柱头直径,花药纵、横径及花丝长度。并拍照。
花粉收集及观察:取六倍体植株即将开放的花序,剥取花药放在洁净的纸盒中,置于室温、干燥、避光处,花药裂开后收集花粉,并于4℃干燥保存备用。将花粉置于蒸馏水中,制成花粉悬液,取少量花粉悬液于载玻片上,加盖玻片,制成临时装片。以四倍体杂种花粉为对照,用Leica DMI6000B荧光显微镜观察花粉形态、测量大小并拍照。
计量单个花药花粉数:随机选取六倍体植株和四倍体杂种植株的5个成熟花药于200μL的离心管中,置于室温、干燥、避光处,打开离心管盖。待花药裂开、花粉爆出后,加入100μL蒸馏水,于振荡器上充分震荡,取悬液于血球计数板上,显微镜下计数。每个材料重复3次,计算均值。
1.2.3 花粉活力的测定
花粉活力的测定采用离体萌发法。参考谢朝添[21]的方法配制烟草花粉离体萌发培养基:0.01%KH2PO4,0.01% CaCl2,0.01% H3BO3和 15% 蔗 糖。25℃培养4 h后显微镜下观察,以花粉管长度大于花粉粒直径作为萌发标准,统计花粉萌发率。

同时观察六倍体、四倍体杂种花、L-8和云烟87的萌发率。
1.2.4 田间杂交试验及种子情况统计
以六倍体、四倍体杂种为母本,以云烟87为父本进行杂交;以云烟87为母本,六倍体、四倍体杂种为父本进行杂交;以四倍体、六倍体为父母本进行自交。于晴朗无风的早上授粉、套袋,每个组合70-100个,观察花粉在柱头上的萌发及花粉管在花柱中的生长情况。15 d后统计座果率,25 d后采集果实,记录种子数,并对种子发芽率进行统计。
花粉管生长情况观察:采集人工授粉48 h的花柱(含柱头)用FAA固定液固定,参照胡适宜[22]的方法对柱头和花柱进行苯胺蓝染色,观察花粉在柱头上的萌发情况和花粉管在花柱中的生长情况。
2 结果与分析
2.1 烟草六倍体植株
L-8四倍体与云烟87八倍体杂交,杂交数7个,获得成熟果实6个,大部分种子均较为饱满。种子萌发生长至5-6叶期时,随机选取50株幼苗进行染色体数目鉴定。结果显示,其中38株为六倍体(2n=6x=72),所占比例为76%,其余为非整倍体,比例为24%。

图1 烟草六倍体植株(A)及其染色体数目(2n=6x=72)(B)
2.2 烟草六倍体花器官特征和花粉形态
烟草六倍体花由5枚雄蕊和1枚雌蕊组成,花丝大多为4长1短,个别为3长2短。与四倍体杂种相比,烟草六倍体花蕾和花药在形态上明显较大(图2-A、B和C,表1)。六倍体单个花粉的数量少于四倍体(图2-D)。对15个花药的花粉量测定结果显示,六倍体单个花药所含花粉量是1.64×104粒,四倍体杂种单个花药所含花粉量是5.90×104粒,二者差异极显著(p<0.01)。按照每朵花有5个花药来计算,烟草六倍体植株每朵花约有8.19×104粒花粉。
在光学显微镜下观察到烟草六倍体花粉形态多样,大小不均一,有椭圆形、圆形等(图2-E-a)。而四倍体杂种花粉大小均一,形态较为一致,均为近圆形(图2-E-b)。经测量六倍体花粉粒直径均值为43.27 μm;四倍体花粉直径均值为36.19 μm,与何川生等[23]的报道一致。

图2 烟草六倍体与四倍体烟草花器官观察
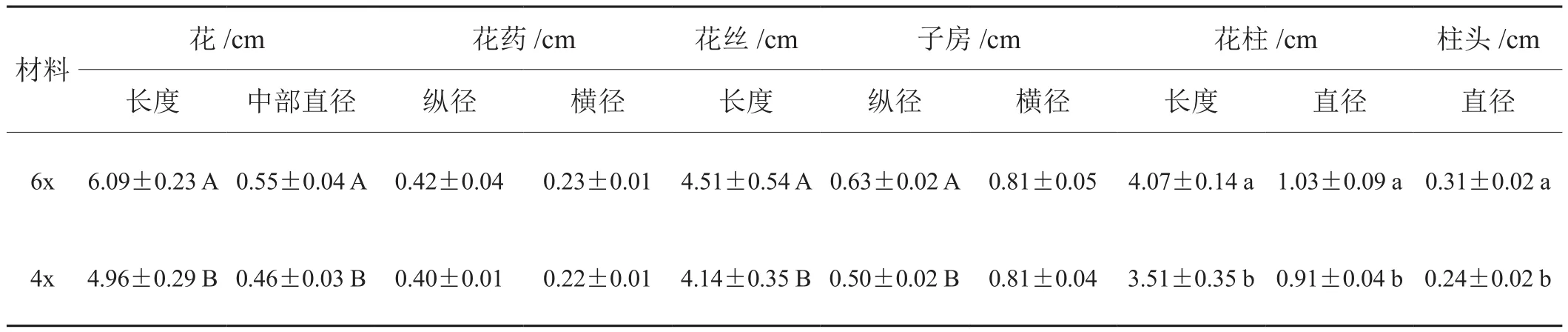
表1 烟草六倍体与四倍体烟草花器官各部特征
2.3 花粉活力测定
从花粉活力测定结果(图3)可以看出,六倍体的花粉和四倍体杂种花粉的活力存在明显差异。六倍体和四倍体杂种的花粉离体萌发率分别为9.78%和57.96%,二者差异极显著(p<0.01)。L-8和云烟87的萌发率分别为76.34%和58.76%。可见,六倍体花粉活力非常低,这可能会造成该六倍体作父本时不育。

图3 烟草六倍体(A)与四倍体杂种(B)花粉的离体萌发
2.4 杂交试验
2.4.1 六倍体、四倍体杂种花粉在柱头上的萌发及生长
六倍体花粉在“云烟87”四倍体植株的柱头上少量萌发(图4-A),明显低于四倍体杂种花粉在云烟87柱头上的萌发率(图4-D)。授以四倍体杂种花粉的云烟87花柱中部及基部大量可见花粉管(图4-E和F),而授以六倍体花粉的云烟87花柱中部和基部只能观察到少量花粉管(图4-B和C)。六倍体自交时,其花粉在柱头上的萌发及在花柱中的生长与其在云烟87柱头上的萌发及花柱中的生长类似,而四倍体杂种自交时花粉的萌发与生长与其做父本同云烟87杂交是类似。可见,六倍体花粉在柱头上的萌发率确较低,且穿柱率也较低,可见六倍体花粉确较低。
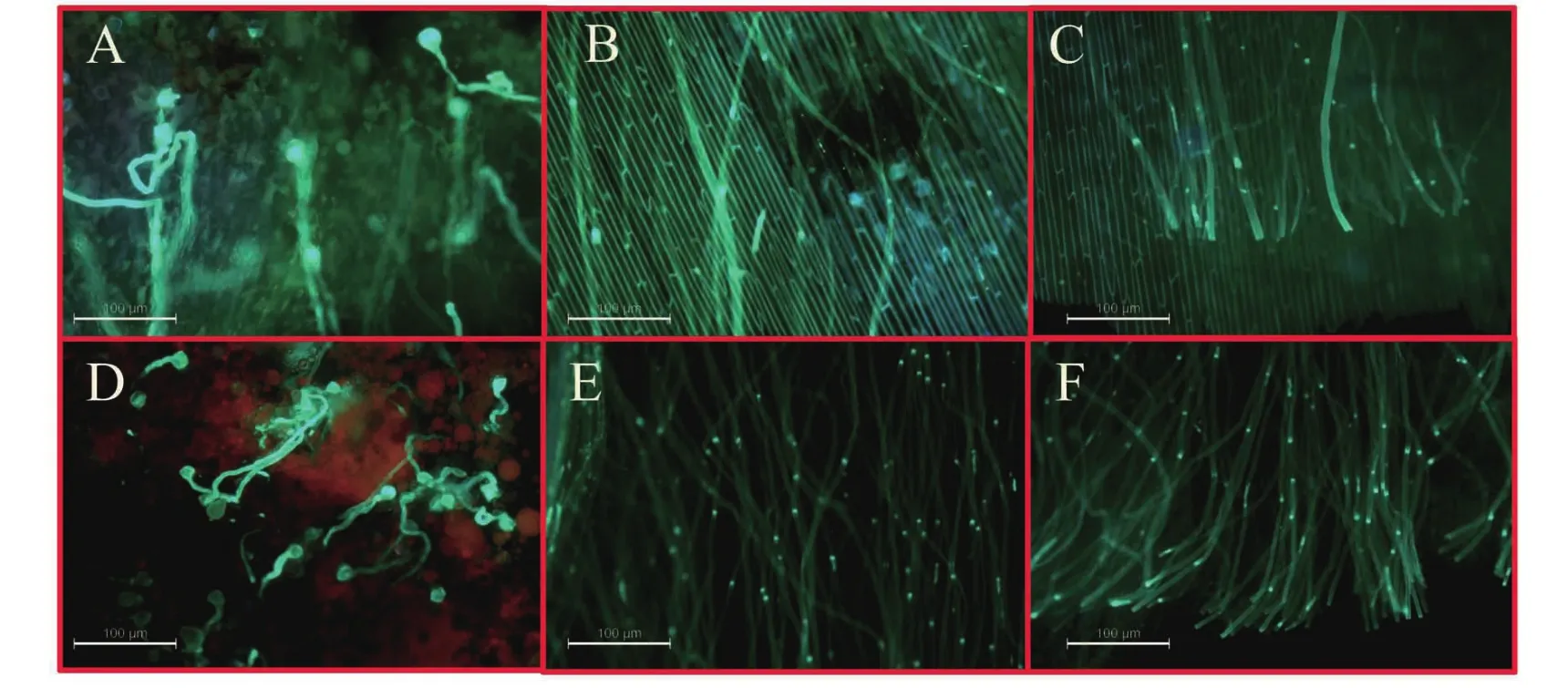
图4 六倍体、四倍体杂种花粉在云烟87四倍体柱头上的萌发及在花柱中的生长
2.4.2 坐果率、种子数量与种子发芽率
授粉15 d后,对杂交各个组合进行统计发现,六倍体自交和以六倍体为父本、云烟87为母本杂交组合的坐果率均为0;以六倍体为母本、云烟87为父本,杂种四倍体为母本、云烟87为父本,杂种四倍体为父本、云烟87为母本三个组合坐果率均为100%。六倍体为父本不能坐果。
以六倍体为母本、云烟87为父本,单果种子数2406.45,千粒重为0.073 g,发芽率为21.2%;杂种四倍体为母本、云烟87为父本,单果种子数为2434.58,千粒重为0.069 g,发芽率为59%;杂种四倍体为父本、云烟87为母本,单果种子数为2508.21,千粒重为0.068 g,发芽率为61.3%。可见六倍体为母本,种子得率与四倍体相近,但其后代大多不能发芽,这可能是六倍体胚死亡所致;六倍体种子较四倍体种子大,且千粒重略大于四倍体,这可能与倍性增加有关。
3 讨论
烟草育种中多采用烟草种内杂交和远缘杂交,前者亲本均为四倍体,后代育性大多较好,后者所获后代多为双单倍体,育性较差,需加倍为双单倍体后方可继续利用[24]。本研究通过烟草八倍体与四倍体杂交获得六倍体,尚属首次。该六倍体具有一定的育性,且亲本之一为对黑胫病具抗性的L-8,因此,该六倍体在后续研究中将有较高的利用价值。
多倍体材料在育种中的重要作用不仅限于其表现出的营养生长优势,还在于其能通过有性生殖产生染色体数量和结构变异的后代[25],如利用大白菜—结球甘蓝[26]、芥蓝—菜薹[27]种间三倍体与亲本之一回交获得异源单体附加系,进而获得易位系材料[28]。利用烟草六倍体与四倍体进行杂交,一方面可以在后代产生众多染色体变异类型;二则因为本试验是通过主栽品种云烟87八倍体与重要的抗病育种材料L-8四倍体进行染色体数量不对称杂交获得的六倍体,因此,在六倍体后代中可以大量得到含云烟87染色体较多而含L-8染色体较少的植株,这在理论上可减少轮回杂交的次数,与传统染色体数量对等的杂交方式相比,有可能缩短育种时间[3]。
众多物种在经染色体数量成倍增加后,其育性均不同程度地降低,其原因是倍性增加导致减数分裂过程中染色体联会异常,终变期形成单价体、三价体及多价体,导致后期正常四分体的数量极少,花粉大多发育不正常,以致其花粉萌发率降低[29-32]。本试验中所得结果与此类似,六倍体花粉萌发率只9.78%,且可少量在柱头上萌发,可见以六倍体为父本时应适当增加授粉数量;作母本时则可正常坐果。
4 结论
虽然六倍体烟草花器官较大,但其花粉量少、形态不均一,且花粉活力较低,这可能是造成其作父本时不能坐果并获得种子的原因之一;六倍体作母本与四倍体杂交可获得一定量的种子,种子萌发率可达21.2%,在后续杂交过程中可重点将其作为母本使用。
[1]许美玲, 张绍芬, 段玉琪,等. 烤烟品种资源抗黑胫病品种综合评价[J].云南农业大学学报, 2003, 18(1): 42-47.
[2]佟道儒. 烟草育种学[M].中国农业出版社,1997:57.
[3]Herrera J C, Combes M C, Cortina H, et al. Gene introgression intoCoffea arabicaby way of triploid hybrids (C. arabica×C.canephora)[J].Heredity, 2002, 89: 488-494.
[4]Lammerts W E. Inheritance of monosomics inNicotiana rustica[J].Genetics, 1932, 17: 689-696.
[5]Clausen R E, Cameron D R. Inheritance inNicotiana tabacum. XVIII. Monosomic analysis [J].Genetics, 1944,29: 447-477.
[6]Cameron D R. The monosomics ofNicotiana tabacum[J].Tob Sci, 1959, 3: 164-166.
[7]Moore G A, Collins G B. Isolation of nullihaploids from diverse genotypes ofNicotiana tabacum[J].J Hered, 1982, 73:192-196.
[8]Reed S M. Cytogenetic evolution and aneuploidy in Nicotiana [M]// Chromosome engineering in plants:genetics, breeding, evolution, Part B, 1991:483-505.
[9]Goodspeed T H. The genusNicotiana. Chronica Botanica[D],Waltham, MA, USA, 1954.
[10] 柴利广, 张俊杰, 林国平,等. 白肋烟两个DH群体的构建与染色体倍性的鉴定[J].中国烟草学报, 2007(2): 33-37.[11] 陈学军, 彭双玉, 罗建蓉,等. 6个烟草杂交组合花药再生苗的培养和DH群体的构建[J].植物遗传资源学报,2011(1): 65-68.
[12]Lewis R S. Transfer of resistance to potato virus Y (PVY)fromNicotiana africanatoNicotiana tabacum: possible in fl uence of tissue culture on the rate of introgression [J].Theor Appl Genet, 2005, 110: 678-687.
[13]Gerstel D U. Inheritance inNicotiana tabacum. XVII.Cytogenetical analysis of glutinosa-type resistance to mosaic disease [J].Genetics, 1943, 28: 553-556.
[14]Gerstel D U. Inheritance inNicotiana tabacum. XIX.Identi fi cation of the tabacum chromosome replaced by one from N. glutinosa in mosaic-resistant Holmes Samsoun tobacco [J].Genetics, 1945, 30: 448-454.
[15]Gerstel D U. Inheritance inNicotiana tabacum. XX. The addition ofNicotiana glutinosachromosomes to tobacco [J].J Hered, 1945, 36: 197-206.
[16]Chen C C, Chen S K, Liu M C, et al. Mapping of DNA markers to arms and sub-arm regions ofNicotiana sylvestrischromosomes using aberrant alien addition lines [J].Theor Appl Genet, 2002, 105: 8-15.
[17]Rao P N, Stoke G W. Cytogenetic analysis of the F1 of haploid×diploid tobacco [J].Genetics 1963, 48: 1423-1433.
[18]魏治中, 魏克强. 烟草远缘杂交[M].北京:中国农业科学技术出版社, 2008: 8-9.
[19]Anna TROJAK-GOLUCH, Apoloniusz BERBEC.Cytological investigations of the interspecific hybrids ofNicotiana tabacumL. ×N. glaucaGrah [J].J Appl Genet,2003, 44(1): 45-54.
[20]Dorota Laskowska, Apoloniusz Berbec. Cytology and fertility of viable hybrids ofNicotiana tabacumL.cv. TB-566 with N.alata Link et Otto [J].J Appl Genet, 2005, 46(1): 11-18.
[21]谢朝添,邱义兰,葛丽丽,等. 烟草花粉萌发和花粉管生长期间柱头和花柱中的钙分布[J].植物生理与分子生物学学报,2005, 31(1):53-61.
[22]胡适宜. 植物胚胎学实验方法(五)检查花粉在柱头上萌发和花粉管在花柱中生长的制片法[J].植物学通报,1994, 11: 58-60.
[23]何川生,卢秀萍,黄学跃,等. 不同烟草品种花粉粒形态的激光共聚焦扫描显微镜观察[J].中国烟草学报,1999,5(1): 30-33.
[24]Dorota Laskowska, Apoloniusz Berbec. Production and characterization of amphihaploid hybrids betweenNicotiana wuttkeiClarkson et Symon andN. tabacumL[J].Euphytica,2012, 183:75-82.
[25] 李云, 张红宇, 汪秀志,等. 水稻同源三倍体( 149-B)×二倍体F2 代稳定群体的验证及分析[J].遗传学报,2004, 31(6): 604- 608.
[26] 刘炜, 申书兴, 王彦华,等. 大白菜—甘蓝异附加系的获得与鉴定. 园艺学报, 2008, 35(2): 207-212.
[27] 任艳蕊, 张成合, 申二巧,等. 芥蓝—菜薹种间三倍体回交子代染色体数鉴定及单体异附加系的选育[J].园艺学报, 2010, 37(2): 213-220.
[28]商少川,王彦华,李晓峰,等. 辐射诱变大白菜—结球甘蓝1 号单体异附加系获得易位系研究[J].中国科技论文在线: http://www.paper.edu.cn.
[29] 彭静, 魏岳荣, 熊兴华. 植物多倍体育种研究进展[J].中国农学通报, 2010, 26(11): 45-49.
[30]万双粉,张蜀宁,张伟,等. 二、四倍体青花菜花粉母细胞减数分裂比较[J].南京农业大学学报, 2007, 30(1): 34-38.[31] 刘文革, 王鸣, 阎志红. 不同倍性西瓜花粉形态观察[J].园艺学报, 2003, 30(3): 328-333.
[32]郭启高,季昆,吴琼,等. 龙泉1号天然三倍体枇杷的花粉形态及生活力[J].果树学报, 2010, 27(3): 391-396.
猜你喜欢
麦类作物学报(2022年8期)2022-08-08
热带作物学报(2022年5期)2022-06-01
中国饲料(2022年5期)2022-04-26
科学养鱼(2021年12期)2022-01-14
东北林业大学学报(2021年12期)2021-02-10
散文诗世界(2019年10期)2019-09-10
蔬菜(2019年5期)2019-05-21
农业科技与装备(2017年9期)2018-03-02
江苏农业科学(2016年8期)2017-02-15
中国果业信息(2015年6期)2015-01-23
