
遮阴对富贵草光合特性的影响1)
2013-03-05 09:58严海燕金光勋马家艳
东北林业大学学报 2013年5期
严海燕 董 然 金光勋 马家艳
(吉林农业大学,长春,130118)
富贵草(Pachysandra terminalis)为黄阳科(Buxaceae)板凳果属(Pachysandra Michx)常绿灌木,高约30 cm,又称转筋草、顶花板凳果、顶蕊三角咪,原产我国湖北、四川、陕西和甘肃等省区,日本也有分布[1]。富贵草作为常绿阔叶植物具有植株低矮、株型美观、枝叶浓密等特点,是我国西北地区优良的乡土地被观赏植物,也是解决园林绿化中强阴下裸露地面覆盖问题和营造冬季常绿景观的优良植物,其带根全草可入药,性苦,微辛,平,具有祛风湿,舒筋活血,通经止带之功效[2]。
目前,国内对富贵草的研究主要集中在化学成分、药理及临床应用等方面,而在光适应性方面只有简单零星的报道。文中主要研究了富贵草在长春地区的引种栽培过程中不同光照条件对其光合特性的影响,旨在掌握富贵草对光环境的适应性,同时为其引种驯化提供科学的理论依据和指导,以期丰富北方地区常绿阔叶树种的多样性和新颖性,优化生态环境。
1 试验地概况
试验在吉林农业大学园艺学院园林苗圃进行。该试验地位于125°24' ~125°36'E、43°47' ~43°61'N,海拔251 m,为大陆性季风气候,春季干燥多风,夏季温热多雨,秋季晴朗早晚温差大,冬季漫长寒冷;年均气温4.9 ℃,最低气温-39.7 ℃,最高气温39.6 ℃,最热月份是7月份,平均气温为23 ℃;年均降水量为565 mm,6—8月份是降水量最丰富时期,占全年降水量的70%;年日照时间2 698 h。
2 材料与方法
于2006年6月份从北京中科院引进富贵草,在吉林农业大学园艺学院园林苗圃进行栽培,每年进行扦插繁殖。2011年5月5日选取健壮且长势一致的富贵草2年生扦插苗定植于20 cm(高)×20 cm(盆底直径)的黑色塑料盆中,栽培基质由V(园土)∶V(菌渣)∶ V(草炭)=2∶ 2∶ 1 混合而成,将盆栽苗移至遮阴棚中缓苗1 个月,期间进行正常的水肥管理。2011年6月5日将盆苗分别移至遮光度不同的黑色遮阴棚(5 m×4 m×3 m,遮阴棚间距3 m,以避免交叉遮光)进行试验处理(用ZDS -10 型数字式照度计测定各处理的透光率):遮阴0、50%、70%、90%,以遮阴0 为对照CK。每处理18 株。
净光合速率日变化的测定:2011年8月初,选择晴朗无云天气,连续3 d 采用Li-Cor6400 便携式光合仪(美国LI - COR 公司生产)从06:00—18:00,每2 h 测定各处理中富贵草的净光合速率(Pn),同时记录蒸腾速率(Tr)、气孔导度(Gs)、胞间CO2摩尔分数(Ci)等相关指标。计算水分利用率(WU,E)=Pn/Tr,气孔限制值(Ls)=1 -Ci/Ca(Ca为大气中CO2摩尔分数)。测定时,每处理选3 株,每株选3 片中、上部功能叶,每叶测3 次。
光合光响应曲线的测定:2011年8月初,采用Li-Cor6400 便携式光合仪(美国LI -COR 公司生产)在0 ~1 400 μmol·m-2·s-1范围内测定Pn,利用仪器的内置LED 红蓝光源按从高到低设置12 个光照梯度:1 400、1 200、1 000、800、600、400、200、150、100、80、50、0 μmol·m-2·s-1,每处理选3 株,每株选3 片中、上部功能叶,每叶测3 次。用叶子飘模型对光合光响应曲线进行拟合[3],叶子飘模型表达式:Pn=a(1 -βⅠ)(Ⅰ-LC,P)/(1 +γⅠ)(其中,α 为光响应曲线的初始斜率;β 为修正系数;γ =α/Pmax;Ⅰ为光照强度)。绘制光合光响应曲线并求出光补偿点(LC,P)、光饱和点(LS,P)、表观量子效率(AQ,Y)、暗呼吸速率(Rd)。采用Excel2003、SPSS17.0 分析软件进行数据处理,采用SigmaPlot10.0 进行作图。
3 结果与分析
3.1 不同遮阴下富贵草光合生理参数日变化情况
3.1.1 不同遮阴下富贵草净光合速率日变化情况
由表1看出,不同遮阴下富贵草叶片的净光合速率均表现出先增加后减少之后再增加最后又减少的变化规律,在12:00 都表现出明显的光合“午休”现象。CK 中富贵草净光合速率的第一个最大值出现在08:00,大小为5.507 μmol·m-2·s-1,第二个最大值出现在16:00,大小为4.372 μmol·m-2·s-1。遮光50%、70%、90%下第一个最大值均出现在10:00,比CK 推迟2 h,大小分别为8.572、9.132、9.971 μmol·m-2·s-1,分别比CK 的第一个最大值高55.66%、65.83%、81.06%。遮光50%的第二个最大值出现在16:00,大小为6.112 μmol·m-2·s-1,与CK 的第二个最大值出现的时间相同,而遮光70%、90%中第二个最大值出现的时间均为14:00,大小分别为7.545、7.932 μmol·m-2·s-1。
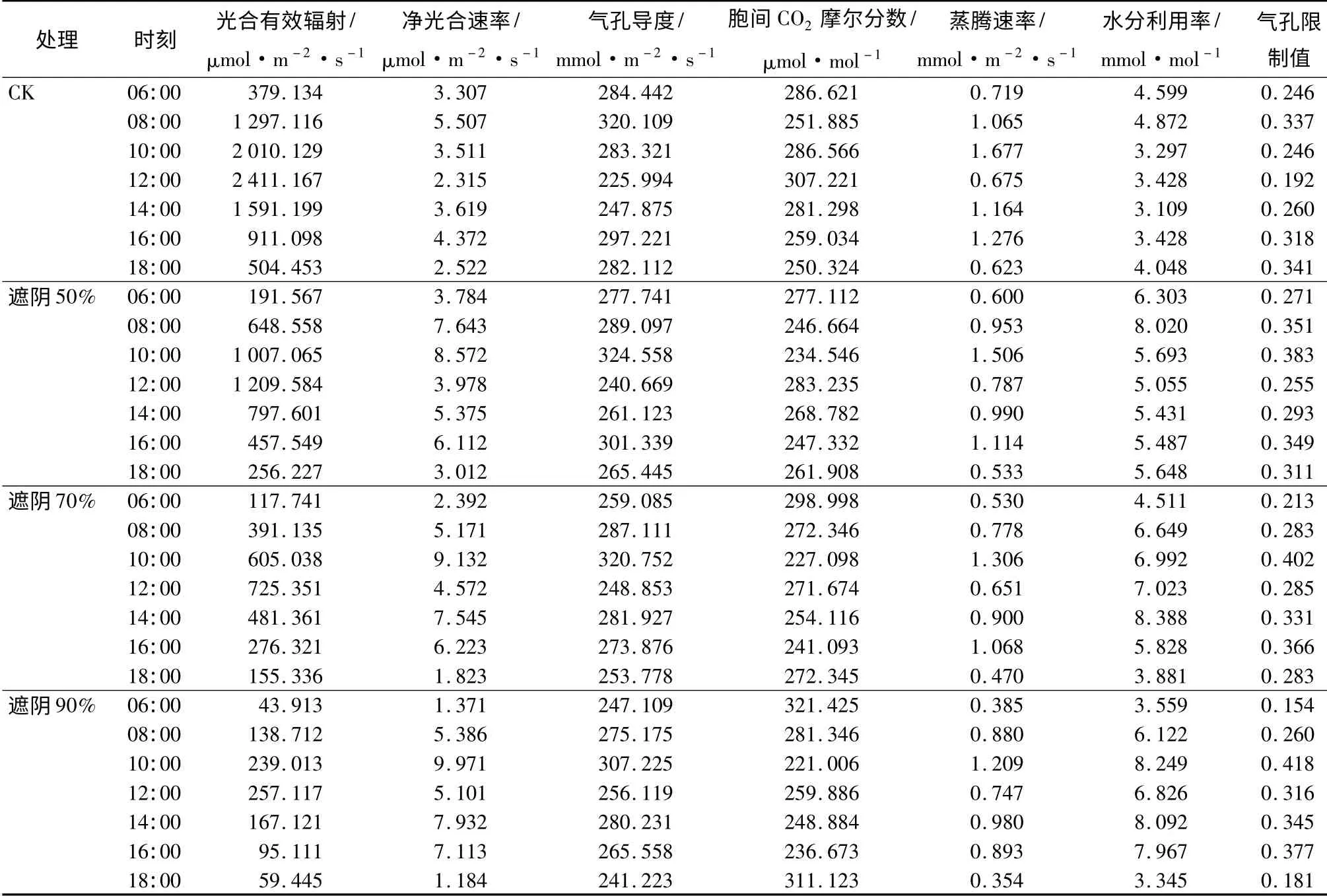
表1 不同遮阴下富贵草光合作用日变化指标比较
由表2可以看出,遮光50%、70%、90%的日均净光合速率依次为5. 497、5. 265、5. 266 μmol·m-2·s-1,分别比CK 的3.593 μmol·m-2·s-1高出52.99%、46.53%、46.56%,但三者之间并没有显著性差异(P >0.05)。
3.1.2 不同遮阴下富贵草气孔导度与胞间CO2摩尔分数日变化情况
由表1可知,不同遮阴下富贵草气孔导度日变化与净光合速率日变化趋势基本一致,在06:00 和18:00,光线较弱,气孔开张小,而在12:00 高温强光使气孔导度降低。不同遮阴下胞间CO2摩尔分数日变化与净光合速率日变化呈现相反的变化规律,12:00 CK 和遮光50%情况下,胞间CO2摩尔分数分别高于各自在06:00 和18:00 的值,说明在CK和遮光50%时富贵草中午利用CO2的能力低于早晚,而遮光70%、90%情况下富贵草中午利用CO2的能力高于早晚。
3.1.3 不同遮阴下富贵草蒸腾速率与水分利用率日变化情况
从表1可知,不同遮阴下富贵草蒸腾速率日变化与净光合速率、气孔导度日变化趋势基本一致,可能因为蒸腾速率受气孔导度的影响较大。不同遮阴下水分利用率日变化并未呈现明显规律,但从表1可以看出,遮光50%、70%、90%中的水分利用率显著高于CK。
3.2 不同遮阴下富贵草的光合光响应曲线及相关参数的变化情况
3.2.1 不同遮阴下富贵草光合光响应曲线的变化情况
由图1知,遮光50%、70%、90%处理下富贵草叶片的净光合速率对光强的响应规律与CK 处理下的基本一致,均有快速增加、饱和与下降阶段:在光强小于200 μmol·m-2·s-1的弱光范围内,随光合有效辐射的增加,净光合速率几乎呈线性增加;在光强大于200 μmol·m-2·s-1而小于各自的光饱和点时,随光合有效辐射的增加净光合速率缓慢增加;当光合有效辐射达到各自的光饱和点时,净光合速率达到最大;当光合有效辐射继续增加,会由于光照过强产生光抑制导致净光合速率下降。从图1可以看出,在产生光抑制后随光强的增加CK 的净光合速率下降速度相对缓慢而遮光90%下降速度最快,且CK 发生光抑制时所需的最小光强明显大于遮光90%的,表明在经过一段时间的处理后遮光90%下生长的富贵草最容易产生光抑制,而生长在CK 下的富贵草由于长期接受全光照的刺激已对强光环境产生一定的适应机制,因此最不容易产生光抑制,这是富贵草长期适应不同光环境的结果。CK 的整个光响应曲线均低于其他3 个处理,可见全光下生长的富贵草光合能力较低。
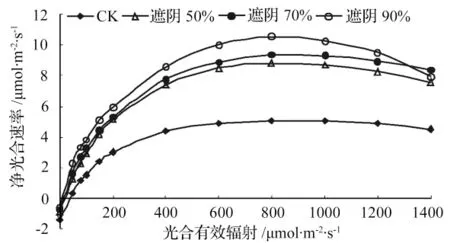
图1 不同遮阴下富贵草叶片的光合光响应曲线
3.2.2 不同遮阴下富贵草光合光响应曲线相关参数的变化情况
遮光对富贵草叶片的最大净光合速率、暗呼吸速率、光补偿点、光饱和点和表观量子效率均有显著的影响(表2)。随着遮光程度的加强,最大净光合速率和表观量子效率均增大,且遮光50%、70%、90%与CK 相比极显著升高(P <0.01);暗呼吸速率、光补偿点和光饱和点均随遮光程度的加强而极显著降低(P <0.01)。其中,遮光90%与CK 相比光补偿点降低76.92%,光饱和点降低39.16%,暗呼吸速率降低46. 62%,表观量子效率增加95.65%,这表明富贵草在弱光环境下,可大幅度降低光补偿点、光饱和点并保持高表观量子效率来提高对弱光的利用能力,同时降低呼吸消耗,以维持植株的正常生长发育,表现出对弱光环境极强的适应能力。最大净光合速率是植物光合能力的体现,遮光50%、70%、90%处理下的富贵草叶片均保持相对较高的最大净光合速率,与CK 相比均极显著升高(P <0.01),表明全光照会大大降低富贵草的光合能力,不适合其生长发育,而在遮光50%、70%、90%的环境中富贵草保持较强的光合能力。
4 结论与讨论
净光合速率日变化常因植物种类和环境条件的不同而表现出差异,同一植物不同部位的叶片在不同月份不同环境条件下可呈现不同的类型[4],通常在自然条件下净光合速率日变化可分为“双峰”型、“单峰”型和“三峰”型。本研究中,不同遮阴下富贵草的净光合速率日变化均表现出先升高后减少再增加,最后减少的变化趋势,属于典型的“双峰”型变化规律,可见遮阴只改变富贵草每日各时刻的净光合速率数值,并未改变净光合速率日变化规律,这与房晓君等[5]的研究结果一致。许大全[6]将植物光合“午休”现象的原因归纳为气孔限制和非气孔限制两大因素,根据Farquhar et al.[7]提出的判断依据,可知富贵草光合“午休”主要是由于非气孔限制因素引起的,这与李金鹏等[8]、赵国禹等[9]的研究结果相似。由日均净光合速率的大小情况可知,富贵草在遮光50%、70%、90%条件下均能保持良好的光合状态,而在全光照下则会出现严重光抑制。

表2 不同遮阴条件下富贵草光响应曲线参数
植物蒸腾速率日变化主要是由于外界环境因子的变化而产生的自身适应性调整,最终呈现出复杂的日变化规律[10],本研究中各遮阴下富贵草的蒸腾速率日变化与净光合速率日变化表现出相似的变化趋势,并且均与气孔导度的变化趋势一致,早晚蒸腾速率低可能是由于低温弱光导致气孔导度下降引起的,而中午蒸腾速率下降则是由于高温强光导致气孔关闭引起的,可见气孔开闭是影响富贵草蒸腾速率的重要因子。
本研究中各遮阴下富贵草水分利用率日变化与净光合速率日变化之间并未呈现明显的线性相关性,这是因为植物水分利用率的大小取决于CO2净同化效率与蒸腾速率,受植物本身根茎叶组织生物结构特征以及大气温度、湿度、气压、土壤水分等环境因子的影响[11]。遮阴50%、70%、90%下的日均水分利用率分别比CK 高55.46%、61.58%、64.90%,说明在遮阴50%、70%、90%下富贵草具有很高的水分利用率,能较好地抵御干旱等逆境。
富贵草的最大净光合速率随着遮阴强度的加大显著增大,这与王建华等[12]对连翘的研究结果恰好相反,原因可能是由于富贵草是喜阴植物,遮阴改善了叶片的光合性能,从而使其最大净光合速率表现出与喜光植物连翘相反的变化。最大净光合速率在遮阴50%、70%、90%的光环境下均较高,而CK 与三者相比却极显著下降(P <0.01),表明富贵草对光环境的适应范围相对较广,而全光照则超出了富贵草的光适应范围。
光补偿点和光饱和点是植物光合生理中2 个重要指标,很大程度上反映了植物对光的利用能力,可用来衡量植物的需光程度,光补偿点低、光饱和点高的植物对光环境的适应性较强[13-14]。通常情况下,灌木的阳生叶最大净光合速率为6 ~20 μmol·m-2·s-1,光补偿点为20 ~50 μmol·m-2·s-1,光饱和点为600 ~1000 μmol·m-2·s-1[15],在遮阴70%、90%条件下富贵草的最大净光合速率、光饱和点均在此范围,但光补偿点则分别为(17 ±1)、(9 ±1)μmol·m-2·s-1,明显小于该范围,说明在遮阴70%、90%中富贵草对弱光的利用能力较一般灌木强,因此,在相对较弱的光下就能进行有机物的正向积累;而CK 中光补偿点和光饱和点均较高,最大净光合速率与其他处理相比则极显著降低(P <0.01),说明富贵草在全光照下会出现严重光抑制现象。
表观量子效率是植物光合作用过程中光能最大转化效率的一种度量,Lee 等[16]指出通常植物的表观量子效率范围是0.02 ~0.05,耐阴植物捕获光量子的能力强,因此具有较高的表观量子效率。本研究中富贵草的表观量子效率随着遮阴度的加强而极显著增大(P <0.01),遮阴90%时达到0.045,说明遮阴使富贵草捕获光量子用于光合作用的能力得到了极大地增强;全光下则仅为0.023,说明全光照会极大地降低表观量子效率不利于富贵草的生长。暗呼吸是植物为维持自身正常代谢而进行物质氧化分解获得能量的过程,植物可以通过降低呼吸速率来适应低光环境[17]。本研究中暗呼吸速率随遮阴度的加大而极显著降低(P <0.01),说明富贵草对弱光具有很好的适应能力,是一种典型的喜阴且极耐阴的常绿灌木。
综上所述,富贵草在全光下与在遮光50%、70%、90%中相比,日均净光合速率、水分利用率、表观量子效率均极显著降低(P <0.01),而光补偿点和暗呼吸速率极显著增加(P <0.01),表明全光照不利于富贵草的生长;在遮光50%、70%、90%的光环境中均保持相对较高的日均净光合速率、表观量子效率和光饱和点,同时暗呼吸速率和光补偿点则较低,表明富贵草喜阴且极耐阴,光适应范围较广。因此,在有一定遮阴环境的小区建筑物周围、林地下均能发挥富贵草作为常绿阔叶小灌木的优势。由于目前对富贵草的研究较少,探索何种光照条件能达到富贵草的耐受极限,还需进一步深入研究,从而为其引种驯化、园林应用提供更有力的理论依据。
[1] 安定国.甘肃省小陇山高等植物志[M]. 兰州:甘肃民族出版社,2002.
[2] 万定荣,钱赬,雷永恕.鄂西土家常用抗风湿类植物药[J]. 中国中药杂志,1993,18(10):581 -584.
[3] 叶子飘.光合作用对光响应新模型及其应用[J].生物数学学报,2008,23(4):710 -716.
[4] 郭宝林,杨俊霞,鲁韧强,等.遮光处理对扶芳藤生长和光合特性的影响[J].园艺学报,2007,34(4):1033 -1036.
[5] 房晓君,王奎玲,刘庆超,等.鹿药耐阴性研究[J].中国农学通报,2011,27(8):79 -82.
[6] 许大全.光合作用气孔限制分析中的一些问题[J].植物生理学通讯,1997,33(4):241 -244.
[7] Farquhar G D,Sharkey T D. Stomatal conductance and photosynthesis[J]. Annual Review Plant Physiology,1982,33:317 -345.
[8] 李金鹏,董然.3 种彩色叶玉簪光合日变化[J]. 东北林业大学学报,2011,39(10):56 -58.
[9] 赵国禹,董然,赫微微. 复序橐吾的光合特性及其影响因子[J].东北林业大学学报,2011,39(1):41 -43.
[10] 董智,马宇飞,李红丽,等.4 个紫花苜蓿品种分枝期光合速率、蒸腾速率日变化及其影响因子分析[J]. 中国草地学报,2009,31(3):67 -71.
[11] 王玉辉,何兴元,周广胜.羊草叶片气孔导度特征及数值模拟[J].应用生态学报,2001,12(4):517 -521.
[12] 王建华,任士福,史宝胜,等. 遮荫对连翘光合特性和叶绿素荧光参数的影响[J].生态学报,2011,31(7):1811 -1817.
[13] 张红瑞,高致明,李志敏.牛膝两个农家品种光合特性及叶片解剖结构的比较[J]. 中国农学通报,2008,24(12):142 -145.
[14] 王刚,孙广玉.不同紫花苜蓿品种光合能力的比较[J]. 东北林业大学学报,2007,35(4):19 -21.
[15] Walter L.植物生态生理学[M].5 版. 翟志习,郭玉海,马永泽,等译.北京:中国农业大学出版社,1997:67 -78.
[16] Lee D W,Bone R A,Tarsis S L,et al. Correlates of leaf optical properties in tropical forest sun and extreme -shade plants[J].America Journal of Botany,1990,77(3):370 -380.
[17] 林波,刘庆.四种亚高山针叶林树种的表型可塑性对不同光照强度的响应[J].生态学报,2008,28(10):4665 -4675.
猜你喜欢
西南农业学报(2021年7期)2021-08-23
节水灌溉(2021年5期)2021-05-26
江苏农业科学(2020年12期)2020-07-20
中国农业科技导报(2020年12期)2020-03-15
西安科技大学学报(社会科学版)(2019年1期)2019-09-10
新课程·下旬(2017年7期)2017-08-14
西北林学院学报(2015年5期)2015-01-03
新课程·中旬(2014年10期)2014-12-26
植物营养与肥料学报(2011年2期)2011-10-26
植物营养与肥料学报(2011年2期)2011-10-26
