
树鼩CD4全长编码序列的克隆及分子特征分析
2012-12-25 06:40田巍威高跃东黄京飞李作生张华堂
Zoological Research 2012年1期
田巍威, 高跃东, 郭 彦, 黄京飞, 肖 昌, 李作生, 张华堂,*
(1. 西南大学 生物技术学院, 重庆 北碚 400715; 2. 云南沃森生物技术股份有限公司, 云南 昆明 650106; 3. 中国科学院昆明动物研究所动物模型与人类疾病机理重点实验室; 免疫生物学实验室, 云南 昆明 650223; 4. 中国科学院昆明生物多样性大型仪器区域中心,云南 昆明 650223; 5. 中国科学院昆明动物研究所 遗传资源与进化国家重点实验室, 云南 昆明 650223)
树鼩CD4全长编码序列的克隆及分子特征分析
田巍威1,2,3, 高跃东4,5, 郭 彦3, 黄京飞4,5, 肖 昌1, 李作生2,*, 张华堂3,*
(1.西南大学 生物技术学院,重庆 北碚400715; 2.云南沃森生物技术股份有限公司,云南 昆明650106; 3.中国科学院昆明动物研究所动物模型与人类疾病机理重点实验室;免疫生物学实验室,云南 昆明650223; 4.中国科学院昆明生物多样性大型仪器区域中心,云南 昆明650223; 5.中国科学院昆明动物研究所 遗传资源与进化国家重点实验室,云南 昆明650223)
树鼩作为多种人类疾病研究模型的可能性已受到广泛关注, 但尚缺乏研究其免疫功能的基本标志以及单克隆抗体。该实验首先以树鼩外周血总RNA为材料, 通过RT-PCR扩增得到长度为1 365 bp的树鼩CD4全长编码序列, 并确定了数据库中缺失的两个片段, 进而通过Clustal W等软件对其序列和分子特征进行分析, 发现树鼩CD4氨基酸序列胞外和胞内域保守性较好,且与人类和猴的亲缘关系较近。虽然树鼩和人CD4分子表面大部分区域均带正电荷, 但与人CD4胞外域D1相比, 树鼩CD4 D1结构区域表面带负电荷较多, 且多出两个N-糖基化位点。这些差异对抗体的结合可能存在影响。该研究为今后树鼩CD4单克隆抗体制备及功能研究奠定了基础。
树鼩; CD4分子; RT-PCR; 结构; 功能; 单克隆抗体
CD4是具有4个Ig样结构域的单链跨膜糖蛋白,主要表达于T淋巴细胞、巨噬细胞、单核细胞, 参与T细胞的活化(Zeitlmann et al, 2001)。作为T辅助细胞(T-helper)活化的协同受体(co-receptor), 其胞外域可以与MHC-II (major histocompatibility complex class II)结合, 稳定TCR与抗原肽-MHC- II复合物之间的相互作用(Grakoui et al, 1999);而其胞内域中的保守基序KKTCQC能与酪氨酸蛋白激酶Src家族成员p56lck结合, 参与T细胞活化的信号转导过程(Glaichenhaus et al, 1991)。
树鼩(tree shrews,Tupaia belangeri)是一类外型酷似松鼠的小型哺乳动物, 主要分布于我国西南部、菲律宾、印度等地(Cao et al, 2003)。早在1987年, Su et al (1987)发现成年树鼩接种人HBV感染血清后, 其血清中能够检测到乙型肝炎表面抗原(hepatitis B surface antigen, HBsAg)。Walter et al (1996)报道新生树鼩经人HBV感染后, 表现出类似人类的急性自限性肝炎。此外, 已有报道树鼩对HCV、HSV亦具有易感性(Zhao et al, 2002; Darai et al, 1980)。由于树鼩对多种人类病毒易感且个体小、易于饲养, 其作为多种人类疾病研究模型的可能性已受到广泛关注。
目前, 针对树鼩CD4的研究较少, 仅有根据树鼩2倍覆盖率基因组结合参考人类CD4 (HGNC ID:1678)预测所得序列, 且存在两处未知片段, 影响了单克隆抗体制备中CD4抗原表位分析。为此,本研究首先以获得树鼩CD4完整编码序列为目的,同时比较分析其分子特征, 为树鼩CD4单克隆抗体的制备和功能研究奠定基础。
1 材料和方法
1.1 动物
本研究采用的滇缅树鼩购于云南省昆明医学院实验动物中心。
1.2 树鼩CD4全长编码序列克隆
根据已有预测树鼩CD4序列(http://asia. ensembl.org/index.html, ENSTBEG00000000662),上游引物设计为:5'-ATGGACCGCCCAGTCCGG-3', 由于上述序列3'端尚不完整, 因此参考人、狨毛猴等物种的CD4序列, 下游引物设计为: 5'-TCACATGAGACTACATGTCTTCTGAAACC-3'(图1)。
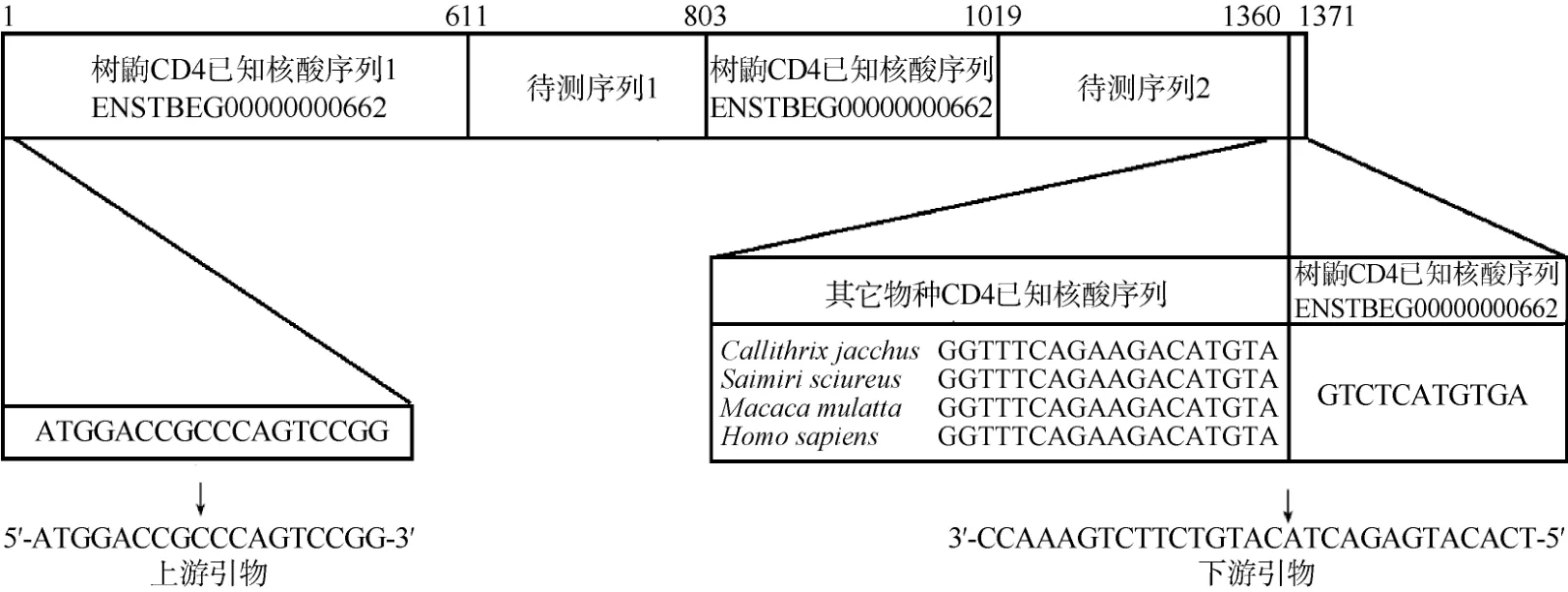
图1 树鼩CD4已知序列和残缺序列以及引物设计Fig.1 Primers for PCR amplification of tree shrews CD4 cDNA
采集树鼩外周血, 提取血液总RNA (试剂盒购于BioTeKe), 将其反转录为cDNA。利用上述引物进行PCR, 循环参数为:94 ℃, 3 min; 94 ℃ 30 s, 45 ℃ 30 s, 72 ℃ 3 min, 32个循环; 72 ℃, 10 min (RT-PCR试剂盒购于TIANGEN)。用树鼩管家基因GAPDH(glyceraldehyde-3-phos-phate dehydrogenase)作为阳性对照。PCR产物经TA克隆插入pMD-19T载体后, 转化大肠杆菌DH5α感受态细胞, 提取质粒进行双酶切鉴定, 最终进行双向测序(北京三博志远基因技术有限公司)。
1.3 分子特征分析
本文用以参比的核酸序列均来源于GenBank,以DNAMAN6.0、BlastP、Clustal W2.0、MEGA4、Discovery Studio和PyMOL分别进行核酸序列、氨基酸序列、物种亲缘关系分析及蛋白质三维结构模建。而信号肽和跨膜域结构分析则采用在线预测(http://www.cbs.dtu.dk/services/SignalP/; http://www. ch.embnet.org/software/TMPRED_form.html)。
2 结果与讨论
2.1 树鼩CD4全长编码序列扩增与比较分析
利用树鼩外周血总RNA, 通过RT-PCR获得长度约为1 365 bp的核酸片段(图2)。通过双向测序,我们确定了所得序列为树鼩CD4分子全长编码序列, 开放阅读框长度为1 365 bp, 编码454个氨基酸(GenBank登陆号为JN657224)。通过DNAMAN 6.0进行多序列比对,所获得树鼩CD4编码序列确定了原预测序列(ENSTBEG00000000662)中第611~797、1 019~1 357的残缺片段, 共计526 bp (图3)。同时, 我们发现已测序列与原预测序列存在差异(表1)。此外, 原预测序列中第798~803、1 358~1 360位9个碱基, 未在我们实测序列中未检出, 而我们实测序列第901~903位的3个碱基在原预测序列中未检出, 但上述差异并未导致移码突变。本实验采用高保真Pfu酶扩增基因并进行两次双向测序, 有效地排除了实验误差。因此, 我们所获得的树鼩CD4分子全长编码序列是较为准确的。
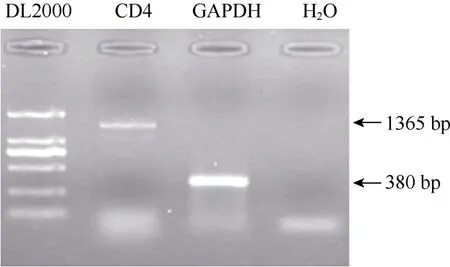
图2 树鼩CD4 cDNA扩增Fig.2 RT-PCR amplification of tree shrews CD4 cDNA

图3 树鼩CD4预测序列未知片段及对应氨基酸Fig.3 The two unknown fragments of the predicted tree shrews CD4 cDNA and the corresponding amino acid
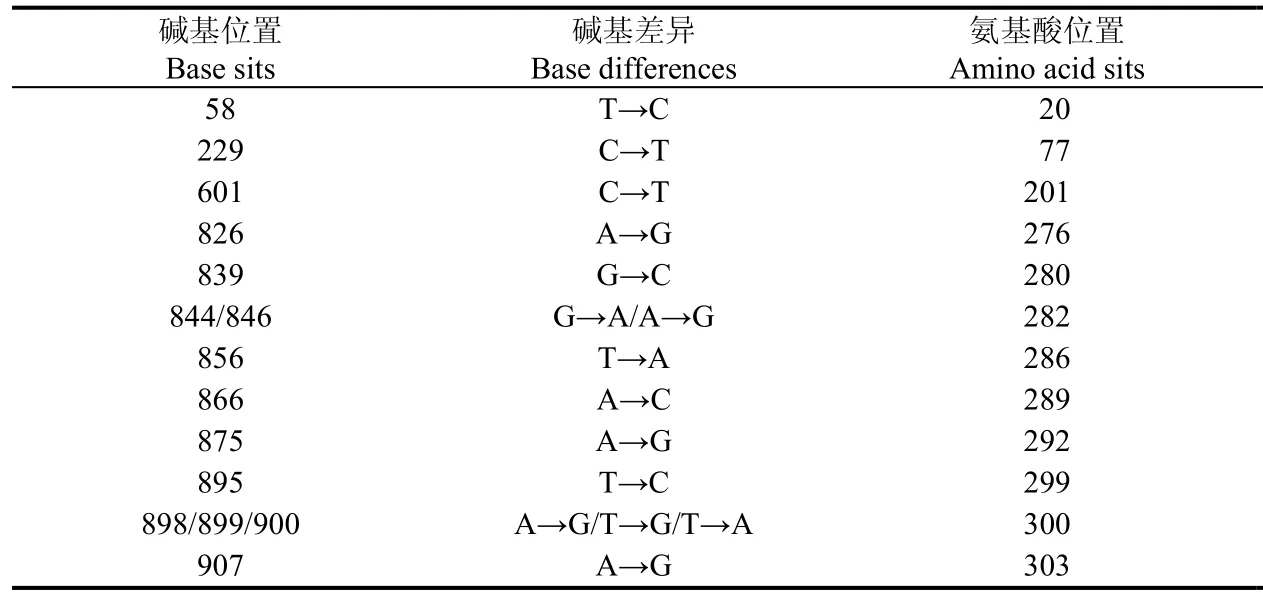
表1 树鼩CD4 cDNA预测序列与实测序列差异性比较Tab. 1 Comparison of the predicted tree shrews CD4 cDNA sequence and the obtained sequence
氨基酸差异
Amino acid differencesSer→ProArg→Trp Ile→Val Gly→Ser Asn→Ser Ser→Thr Thr→Ala Gln→Arg Val→Gly Ser→Thr Thr→Leu Arg→Gly
2.2 树鼩CD4氨基酸序列分析
2.2.1 树鼩CD4整体结构与亲缘关系分析

图4 树鼩CD4推导氨基酸序列比对分析Fig.4 Alignment of deduced amino acid sequences of tree shrews CD4 with those from other mammals
树鼩CD4分子由454个氨基酸组成, 属于Ⅰ型跨膜蛋白。信号肽和跨膜域在线预测显示, 树鼩CD4整体结构与人CD4相似, 可分为信号肽(1~25)、胞外域(26~392)、跨膜域(393~416)、胞内域(416~454)四部分(图4), 我们采用Blast P及Clustal W2.0对推导得到的树鼩CD4(JN_657224)氨基酸序列与人(NM_000616)、恒河猴(NM_001042662)、狼犬(NM_001003252)、穴兔(NM_001082313)、绵羊(NM_001129902)、大鼠(NM_013488)、小鼠(NM_012705)等哺乳动物进行比较(图4), 发现树鼩CD4氨基酸序列与人类、恒河猴的相似度较高, 分别为61%、60%, 而与大鼠、小鼠的相似度较低, 分别为48%、46%。同时, 我们利用MEGA4对树鼩CD4亲缘关系进行分析, 结果显示, 树鼩与人类、恒河猴聚集, 而与大鼠、小鼠距离较远, 说明树鼩在进化关系上跟人类和恒河猴更接近(图5)。
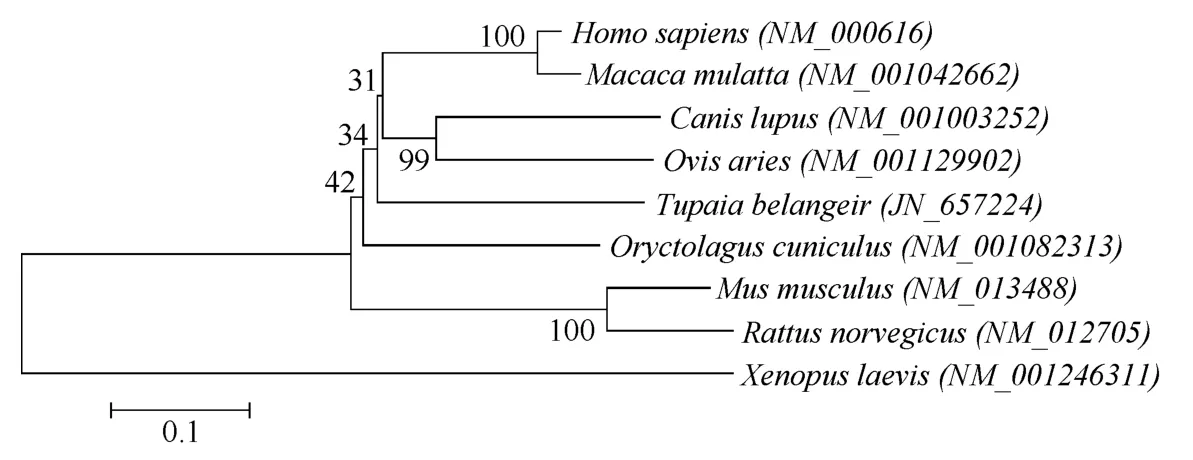
图5 树鼩和其他哺乳动物CD4之间的亲缘关系Fig.5 Phylogentic relationships between CD4 chains from tree shrews and other mammals
2.2.2 树鼩CD4胞外域特征
人CD4胞外域主要由4个免疫球蛋白样结构D1、D2、D3、D4组成, 胞外域中存在3对二硫键。它们分别是形成于D1结构的Cys41和Cys109;D2结构的Cys155和Cys184;D4结构的Cys328和Cys370(Matthias et al, 2002)。通过Discovery Studio,我们对人和树鼩的CD4胞外域进行了三维空间结构分析。结果显示, 树鼩CD4胞外域和人CD4胞外域整体相似, 都可分为四个免疫球蛋白样结构,在树鼩的D1和D4结构域中能找到相应的二硫键,但形成D2中二硫键的Cys155未能找到, 相应位置被色氨酸占据(图6B)。在CD4氨基酸多序列比对中, 狼犬、穴兔、绵羊的CD4胞外域相应位置同样也为色氨酸(图4)。由于这种差异并没有破坏树鼩D2结构的形成, 我们认为色氨酸的存在可能在一定程度上弥补了二硫键缺失所带来的影响, 但这需要进一步实验验证。
同时, 人CD4分子中保守性氨基酸残基Leu134、Leu202、Leu225、Leu308能够形成疏水域, 而Gln137和Gly306之间能够形成分子内氢键, 它们能够保证D1和D2之间以及D3和D4之间的紧密连接(Wang et al, 1990)。在树鼩CD4胞外域的相应位置同样存在上述保守性氨基酸, 这些氨基酸在维持树鼩CD4稳定性方面可能起到重要作用。进一步分析发现, 人D1中的α-螺旋在树鼩中同样存在,而α-螺旋的存在一定程度可以确保肽链的正确折叠和空间结构的形成。综合以上分析, 我们所获得的树鼩CD4分子胞外域能够形成同人相似的三维空间结构。
人CD4胞外区与MHC-II的结合主要依靠D1、D2结构,特别是D1上的4个带正电的氨基酸残基(Lys60、Phe68、Lys71和Arg84)(Wang et al, 2001),但在我们的序列比对结果中发现, 树鼩CD4分子D1中的后3个核心氨基酸残基都与人存在差异, 而这种差异在氨基酸多序列比对的其它物种中同样存在(图4)。这说明在各物种间CD4与MHC-II结合的氨基酸位点并不保守。CD4的D1结构同时也是大多数单克隆抗体结合的区域, 如BD公司针对人CD4D1区的鼠抗人CD4抗体(克隆号:RPA-T4)。通过人与树鼩CD4胞外域三维空间结构比较, 我们发现树鼩在D1区存在两个N-糖基化位点, 而在人的D1区却不存在(图6A, B), 在人CD4 D3中存在一个N-糖基化位点, 在树鼩CD4分子的D3中未检测到, 而在树鼩CD4分子的D4中包含一个人D4结构中不存在的α-螺旋(图6C, D)。以上差异并没有导致树鼩与人CD4胞外域产生较大不同, 其机制有待进一步探讨。此外, 我们对人与树鼩CD4胞外域的绝对电荷分布进行分析, 发现人和树鼩CD4表面大部分区域均带正电荷, 负电荷主要分布于D1和D4表面, 且树鼩D1表面负电荷分布较人D1表面多(图6E, F)。树鼩CD4的D1区在N-糖基化位点数目和表面电荷上与人CD4的差异可能影响树鼩CD4分子与其它分子的结合, 例如我们在实验中发现鼠抗人CD4抗体不能与树鼩CD4发生交叉反应。
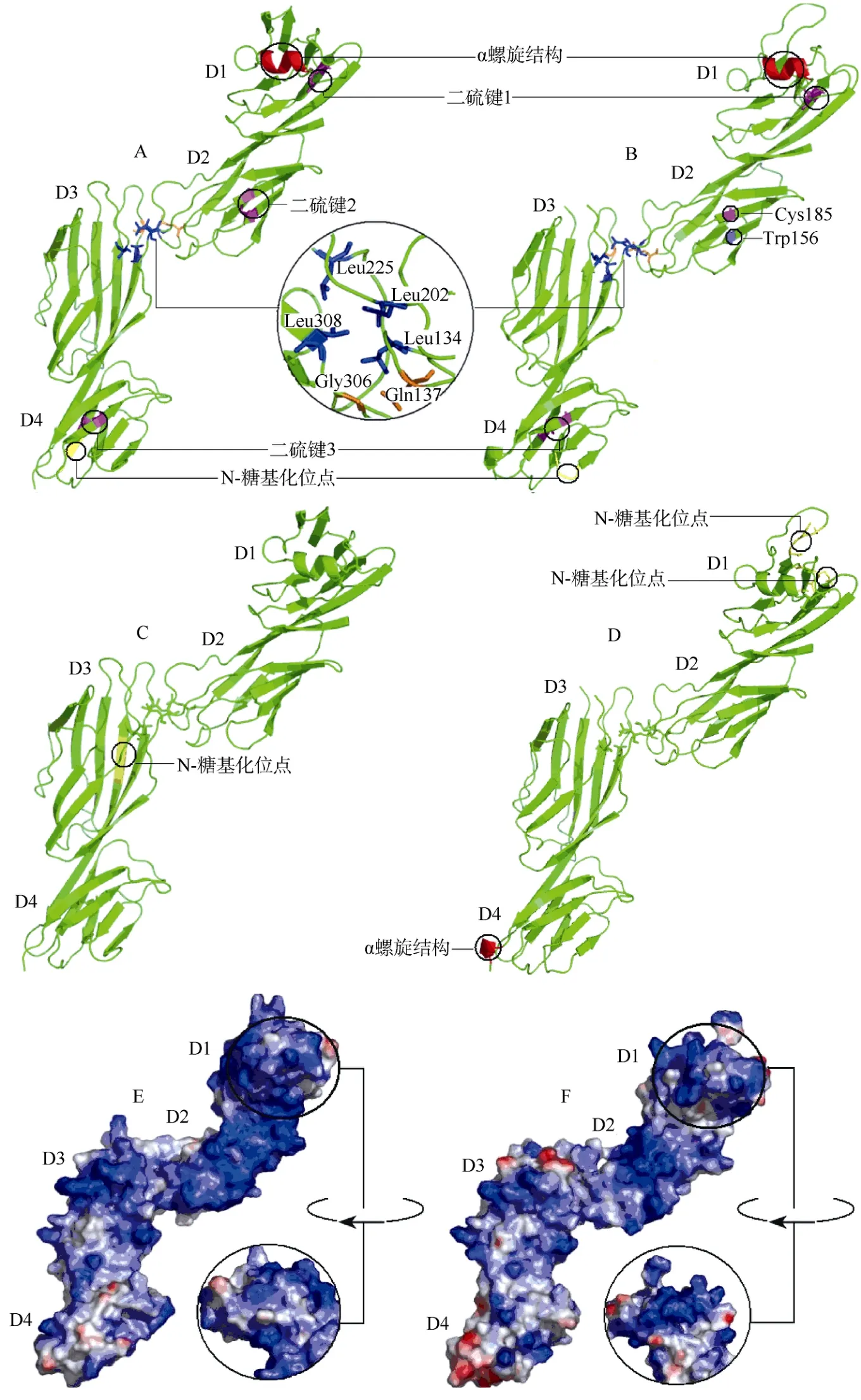
图6 人和树鼩CD4胞外域蛋白三维结构模建和表面电荷比较Fig.6 Modeling of three-dimensional structures and surface charges of the extracellular domain of human and tree shrews CD4 chains
2.2.3 树鼩CD4胞内域特征
CD4氨基酸多序列比对显示树鼩和其它物种的CD4胞内域高度保守(图4), 提示该结构域参与了CD4分子重要功能。Glaichenhaus et al (1991)报道CD4分子胞内域中KKTCQC基序是Src家族酪氨酸蛋白激酶p56lck的结合区域, 其中的两个半胱氨酸是CD4与p56结合所必需的(Learmont et al, 1999),而p56lck在去磷酸化激活状态下, 能够造成CD3胞内域ITAM的磷酸化, 进而引起fyn和ZAP-70等下游信号分子的激活, 最终完成T细胞活化(Leo & Schraven, 2001; Kane et al, 2000)。在树鼩CD4胞内域中, 我们同样检测到高度保守的KKTCQC基序的存在(图4), 明确提示树鼩CD4具有与p56lck结合的结构基础, 进而参与树鼩T淋巴细胞的活化。
我们在树鼩KKTCQC基序上游检测到两个保守的丝氨酸残基和两个保守的亮氨酸残基(图4)。这4个保守氨基酸参与人CD4的内化(internalization),其中两个保守的丝氨酸残基的磷酸化是CD4内化的信号, 而CD4分子内化可以下调细胞表面CD4的表达, 控制T淋巴细胞的过度活化, 保持免疫系统的平衡(Shin et al, 1990)。我们推测树鼩CD4胞内域中的这4个保守性氨基酸亦参与了树鼩CD4的内化过程。此外, 我们发现以上基序在比对的其他哺乳动物中也同样存在(图4), 提示大多数哺乳动物CD4胞内域重要基序在长期进化过程中具有高度的保守性。
综上, 本研究通过RT-PCR扩增得到树鼩CD4全长编码序列, 确定了原预测序列中未知的两处片段, 同时, 对其分子特征进行了比较分析。结果表明:树鼩CD4氨基酸序列与人和恒河猴的亲缘关系较近, 其胞外域整体结构与人CD4胞外域相似的。而CD4与MHC-II结合相关的4个核心氨基酸在各物种中差异较大, 同时, 胞外域的潜在N-糖基化位点、α-螺旋以及表面电荷的分布也存在差异。这些差异的存在有利于对树鼩CD4结构与功能特点的进一步分析。
Cao J, Yang EB, Su JJ, Li Y, Chow P. 2003. The tree shrews: Adjuncts and alternatives to primates as models for biomedical research[J]. J Med Primatol,32(3): 123-130.
Darai G, Zöller L, Matz B, Schwaier A, Flügel RM, Munk K. 1980. Experimental infection and the state of viral latency of adult tupaia with herpes simplex virus type 1 and 2 and infection of juvenile tupaia with temperature-sensitive mutants of HSV Type 2 [J]. Arch Virol,65(3-4): 311-318.
Glaichenhaus N, Shastri N, Littman DR, Turner JM. 1991. Requirement for association of p56lck with CD4 in antigen-specific signal transduction in T cells[J]. Cell,64(3): 511-520.
Grakoui A, Bromley SK, Sumen C, Davis MM, Shaw AS, Allen PM, Dustin ML. 1999. The immunological synapse: a molecular machine controlling T cell activation[J]. Science,285(5425): 221-227.
Kane LP, Lin J, Weiss A. 2000. Signal transduction by the TCR for antigen[J]. Curr Opin Immunol,12(3): 242-249.
Learmont JC, Geczy AF, Mills J, Ashton LJ, Raynes-Greenow CH, Garsia RJ, Dyer WB, McIntyre L, Oelrichs RB, Rhodes DI, Deacon NJ, Sullivan JS. 1999. Immunologic and virologic status after 14 to 18 years of infection with an attenuated strain of HIV-1. A report from the Sydney Blood Bank Cohort[J]. N Engl J Med,340(22): 1715-1722.
Leo A and Schraven B. 2001. Adapters in lymphocyte signalling[J]. Curr Opin Immunol,13(3): 307-316.
Matthias LJ, Yam PTW, Jiang XM, Vandegraaff N, Li P, Poumbourios P, Donoghue N, Hogg PJ. 2002. Disulfide exchange in domain 2 of CD4 is required for entry of HIV-1[J]. Nat Immunol,3(8): 727-732.
Shin J, Doyle C, Yang Z, Kappes D, Strominger JL. 1990. Structural features of the cytoplasmic region of CD4 required for internalization[J]. EMBO J,9(2): 425-434.
Su JJ, Yan RQ, Gan YQ, Zhou DN, Huang DR, Huang GH. 1987. Adult tree shrews experimentally infected with hepatitis B virus[J]. Chn J Pathol,16(2): 103-105. [苏建家,严瑞琪,甘友全,周德南,黄定瑞,黄国华. 1987.成年树鼩实验感染乙型肝炎病毒的研究[J].中华病理学杂志,16(2): 103-105.]
Walter E, Keist R, Niederost B, Pult I, Blum HE. 1996. Hepatitis B virus infection of tupaia hepatocytes in vitro and in vivo[J]. Hepatology,24(1): 1-5.
Wang JH, Meijers R, Xiong Y, Liu JH, Sakihama T, Zhang R, Joachimiak A, Reinherz EL. 2001. Crystal structure of the human CD4 N-terminal two-domain fragment complexed to a class II MHC molecule[J]. Proc Natl Acad Sci USA,98(19): 10799-10804.
Wang JH, Yan YW, Garrett TP, Liu JH, Rodgers DW, Garlick RL, Tarr GE, Husain Y, Reinherz EL, Harrison SC. 1990. Atomic structure of a fragment of human CD4 containing two immunoglobulin-like domains[J]. Nature,348(6300): 411-418.
Zhao X, Tang ZY, Klumpp B, Wolff-Vorbeck G, Barth H, Levy S, von Weizsäcker F, Blum HE, Baumert TF. 2002. Primary hepatocytes of Tupaia belangeri as a potential model for hepatitis C virus infection [J]. J Clin Invest,109(2): 221-232.
Zeitlmann L, Sirim P, Kremmer E, Kolanus W. 2001. Cloning of ACP33 as a novel intracellular ligand of CD4[J]. J Biol Chem,276(12): 9123-9132.
Cloning of full-length coding sequence of tree shrew CD4 and prediction of its molecular characteristics
TIAN Wei-Wei1,2,3, GAO Yue-Dong4,5, GUO Yan3, HUANG Jing-Fei4,5, XIAO Chang1, LI Zuo-Sheng2,*, ZHANG Hua-Tang3,*
(1. College of Biotechnology, Southwestern University, Beibei 400715, China; 2. Yunnan Walvax Biotechnology Co., LTD Kunming 650106, China; 3. Immunobiology Laboratory, Key Laboratory of Animal Models and Human Disease Mechanisms, Kunming Institute of Zoology, the Chinese Academy of Science, Kunming 650223, China; 4. Kunming Biological Diversity Regional Center of Large Apparatus and Equipment, the Chinese Academy of Sciences, Kunming 650223, China; 5. State Key Laboratory of Genetic Resources and Evolution, Kunming Institute of Zoology, the Chinese Academy of Sciences, Kunming 650223, China)
The tree shrews, as an ideal animal model receiving extensive attentions to human disease research, demands essential research tools, in particular cellular markers and monoclonal antibodies for immunological studies. In this paper, a 1 365 bp of the full-length CD4 cDNA encoding sequence was cloned from total RNA in peripheral blood of tree shrews, the sequence completes two unknown fragment gaps of tree shrews predicted CD4 cDNA in the GenBank database, and its molecular characteristics were analyzed compared with other mammals by using biology software such as Clustal W2.0 and so forth. The results showed that the extracellular and intracellular domains of tree shrews CD4 amino acid sequence are conserved. The tree shrews CD4 amino acid sequence showed a close genetic relationship withHomo sapiensandMacaca mulatta. Most regions of the tree shrews CD4 molecule surface showed positive charges as humans. However, compared with CD4 extracellular domain D1 of human, CD4 D1 surface of tree shrews showed more negative charges, and more two N-glycosylation sites, which may affect antibody binding. This study provides a theoretical basis for the preparation and functional studies of CD4 monoclonal antibody.
Tree shrews; CD4; RT-PCR; Structure; Function; Monoclonal antibody
田巍威, 硕士研究生, E-mail: vivit1986@126.com
Q959.832; Q951.3; Q347
A
0254-5853-(2012)01-0060-07
10.3724/SP.J.1141.2012.01060
2011-10-09;接受日期:2012-01-05
国家重点基础研究发展计划(973计划)分课题(2007CB512807);中科院知识创新工程重要方向项目(KSCX2-EW-R-12);树鼩基础生物学及重要生物学特性研究(Y002732073);中科院昆明动物研究所“动物模型与人类疾病机理重点实验室”开放课题
∗通信作者(Corresponding authors),E-mail: zhanght@post.kiz.ac.cn; jxdxlzs@yahoo.com.cn
猜你喜欢
云南画报(2022年3期)2022-04-19
农村青少年科学探究(2020年5期)2020-08-18
民族音乐(2019年2期)2019-12-10
中国洗涤用品工业(2019年4期)2019-05-11
中国(韩文)(2018年6期)2018-06-25
新民周刊(2018年8期)2018-03-02
中成药(2018年1期)2018-02-02
饮食科学(2017年12期)2018-01-02
中成药(2017年3期)2017-05-17
少儿科学周刊·少年版(2015年1期)2015-07-07
