
脂氧素A4抑制内毒素诱导的人支气管上皮细胞环氧合酶2及前列腺素E2的表达*
2012-11-06 05:22朱天琦郑声星金胜威
中国病理生理杂志 2012年3期
朱天琦, 郑声星, 金胜威
(温州医学院1附属第一医院麻醉科,2附属第二医院麻醉科,3附属第二医院ICU,浙江 温州 325000)
1000-4718(2012)03-0478-05
2011-07-09
2011-12-09
国家自然科学基金资助资助项目(No.81070061);浙江省自然科学基金项目(No.Y2100885);浙江省卫生高层次创新人才培养项目
△通讯作者 Tel: 0577-88879006; E-mail: jinshengwei69@yahoo.com.cn
脂氧素A4抑制内毒素诱导的人支气管上皮细胞环氧合酶2及前列腺素E2的表达*
朱天琦1, 郑声星2, 金胜威3△
(温州医学院1附属第一医院麻醉科,2附属第二医院麻醉科,3附属第二医院ICU,浙江 温州 325000)
目的探讨脂氧素A4对人支气管上皮细胞(HBECs)环氧合酶2(COX-2)表达的影响。方法应用不同浓度(0.1、1、10 mg/L)的内毒素(LPS)刺激HBECs 9 h,或者用1 mg/L LPS分别刺激HBECs不同时点(3 h、6 h、9 h)后,测定HBECs的COX-2 mRNA表达和细胞上清液前列腺素E2(PGE2)水平。应用不同浓度 (0、100、400 μmol/L) 的脂氧素A4作用于经过LPS(1 mg/L)刺激培养9 h的HBECs,采用酶联免疫吸附法(ELISA)检测细胞上清液PGE2的水平, 同时分别应用RT-PCR和Western blotting分别检测HBECs COX-2 mRNA及蛋白的表达。结果LPS刺激培养条件下HBECs的COX-2 mRNA表达及其上清液PGE2水平增加,并呈时间、剂量依赖性。脂氧素A4能抑制LPS刺激培养HBECs COX-2蛋白和mRNA的表达及上清液PGE2的水平,并呈剂量依赖性。结论脂氧素A4能抑制LPS诱导的HBECs COX-2表达及上清液PGE2的水平。
脂氧素A4; 人支气管上皮细胞; 环氧合酶2; 脂多糖类
急性呼吸窘迫综合征(acute respiratory distress syndrome,ARDS)是临床常见危重病,其病死率达35%~58%[1],内毒素血症引起的ARDS极为常见。传统观念认为急性炎症效应细胞主要有白细胞及巨噬细胞等,目前越来越多证据表明人支气管上皮细胞(human bronchial epithelial cells, HBECs)不仅作为构成外部环境与机体内环境相互作用的一道屏障,也可以作为急性炎症效应细胞[2]。在呼吸道炎症疾病中人支气管上皮细胞能分泌炎症介质及细胞因子如前列腺素E2(prostaglandin E2,PGE2)、白细胞介素-8(interleukin-8,IL-8)等,同时能表达环氧合酶2(cyclooxygenase 2,COX-2)[3]。COX-2 是合成前列腺素的关键限速酶, 在急性炎症时受多种炎症介质(如细胞因子)刺激而大量表达[3-4]。脂氧素(lipoxins, LXs)是一类重要的内源性脂质抗炎介质,是花生四烯酸(arachidonic acid, AA)的脂加氧酶(lipoxygenase)代谢产物, 由5-脂加氧酶(5-lipoxygenase)与15-脂加氧酶(15-lipoxygenase)或者由5-脂加氧酶(5-lipoxygenase)与12-脂加氧酶(12-lipoxygenase)作用而形成[5],主要通过跨细胞途径来合成 ,在炎症反应中发挥广泛的抗炎促炎症消退作用[6]。本文通过对人支气管上皮细胞在内毒素直接刺激下其环氧合酶-2及PGE2表达和脂氧素A4(LXA4)对人支气管上皮细胞COX-2及PGE2表达的影响进行研究,阐明脂氧素A4在抑制支气管上皮细胞炎症反应中的机制和作用。
材 料 和 方 法
1材料
1.1细胞株 人支气管上皮细胞(武汉大学保藏中心)。
1.2主要试剂 新生牛血清、DMEM培养基均购自Gibco,脂多糖(lipopolysaccharide,LPS,E.coliserotype O55∶B5)及琼脂糖(Sigma), Trizol( Invitrogen),逆转录试剂盒(Fermentas),TAG酶及dNTP (Tiangen),二乙基焦碳酸酯 (DEPC,武汉博士德生物工程有限公司), COX-2引物及阳性对照GAPDH引物(上海赛百盛基因技术有限公司合成),兔抗人COX-2抗体(Abcam),辣根酶标记山羊抗兔IgG(北京中杉公司),PGE2ELISA试剂盒(R&D), BCA蛋白定量试剂盒(Pierce)。
2方法
2.1细胞培养及实验分组 人支气管上皮细胞株常规培养于10%新生牛血清、青霉素1×105IU/L、链霉素1×105IU/L的 DMEM培养基,待细胞呈融合状态时,无血清 DMEM培养基继续培养24 h。实验分4组: (1)正常对照组:不给予任何处理;(2)9 h LPS处理组:同一时点(9 h)给予不同浓度 LPS(0.1 mg/L、1 mg/L、10 mg/L);(3)1 mg/L LPS处理组:不同时点(3 h、6 h、9 h)给予同一浓度LPS(1 mg/L)刺激(3 h、6 h、9 h);(4)脂氧素A4及LPS处理组:给予不同浓度(0 μmol/L、100 μmol/L、400 μmol/L)脂氧素A4加上LPS(1 mg/L)刺激9 h。
2.2ELISA法检测细胞上清液中PGE2的含量 LPS及脂氧素A4作用结束后收集细胞上清液,2 000 r/min离心 5 min后待测。按照R&D公司ELISA试剂盒说明书操作,检测细胞上清液中PGE2的含量。
2.3半定量 RT- PCR检测 COX-2 mRNA的表达 取不同实验组的人支气管上皮细胞,采用Trizol试剂,参照说明书对各实验组细胞的总 RNA进行提取。取1 μg RNA逆转录合成 cDNA,其中1 μL cDNA为模板,扩增 COX-2。反应条件:94 ℃ 预变性5 min, 94 ℃ 45 s,56 ℃ 40 s,72 ℃ 3 min,35个循环,最后72 ℃ 7 min。取 5 μL扩增产物用1.5%琼脂糖凝胶电泳,溴化乙啶 (EB) 显色,目的条带用图像分析系统做灰度值测定,以GAPDH为内参照,COX-2与GAPDH灰度值比值作为COX-2 mRNA的相对含量进行半定量分析。COX-2的引物序列:上游引物5’-AAGCTGGGAAGCCTTCTCTA-3’,下游引物5’-TTTCCATCCTTGAAAAGGCGC-3’ ,扩增片段长度为342 bp。GAPDH的引物序列:上游引物 5’-ACCAGCCCCAGCAAGAGCACAAG-3’,下游引物5’- TTCAAGGGGTCTACATGGCAACTG -3’,扩增片段长度为123 bp,反应条件: 94 ℃ 预变性5 min,94 ℃ 45 s,56 ℃ 40 s,72 ℃ 3 min。
2.4Western blotting检测COX-2蛋白的表达 取1 mg/L LPS合并不同浓度LXA4诱导培养9 h后的支气管上皮细胞,加入适量细胞裂解液,冰上孵育 40 min,12 000 r/min离心20 min,上清液即为总蛋白,参照BCA蛋白定量试剂盒说明书测蛋白浓度。将总蛋白进行SDS PAGE凝胶电泳,然后电转移至NC膜上。用5%脱脂牛奶对膜室温封闭1 h,加入兔抗人COX-2和小鼠抗人β-actin多克隆抗体(COX-2以 1∶500稀释,β-actin以 1∶2 000稀释 ) 4 ℃摇动过夜。第2 d用 TBST于室温下洗膜3次,每次15 min,接着加入辣根酶标记山羊抗兔IgG抗体(1∶2 000稀释 )及山羊抗小鼠IgG抗体(1∶2 000稀释 ),在室温下轻轻摇动1 h,然后用TBST室温下洗膜 3次,每次15 min。最后用化学发光法曝光显迹,按化学发光显影试剂盒 (ECL)说明要求加入ECL试剂,室温下温育 2 min后对 X线片曝光。
3统计学处理
结 果
1LPS刺激下人支气管上皮细胞COX-2mRNA的表达及其上清液PGE2含量呈剂量、时间依赖关系
通过RT-PCR检测发现,正常人支气管上皮细胞中COX-2 mRNA有微量表达,LPS诱导后支气管上皮细胞COX-2 mRNA表达增加,与对照组相比有差异显著(P< 0.01);用不同浓度 LPS(0.1、1和10 mg/L) 刺激细胞9 h后, COX-2 mRNA表达亦随之增多,1 mg/L、10 mg/L组 LPS刺激COX-2 mRNA表达达到高峰,两者间差异无统计学意义,与0.1 mg/L组相比差异有统计学意义(P<0.01),见图1A,1B。用1 mg/L LPS刺激培养HBECs后,在不同时点(3 h、6 h、9 h)COX-2 mRNA表达亦增多, 并呈时间依赖关系,9 h组与3 h、6 h组相比显著差异(P< 0.01),结果见图1A、C。mRNA 表达的半定量结果以COX-2/β- actin吸光度的相对值计算。而且用LPS刺激HBECs能增加上清液PGE2(PGE2是COX-2主要产物之一)的含量,呈剂量、时间依赖关系。LPS诱导后,各组与对照组相比差异显著(P< 0.01),1 mg/L、10 mg/L 组PGE2含量分别为(92.606±1.977)ng/L和(92.716±2.947)ng/L,与0.1 mg/L组[(46.332 ±1.609)ng/L相比有显著差异(P<0.01),3 h和6 h组PGE2含量分别为(47.501±1.527)ng/L和(60.918±1.122)ng/L,与9 h组[(91.966±3.246)ng/L]相比有差异显著(P<0.01),见图2。
2脂氧素A4对LPS刺激的人支气管上皮细胞COX-2mRNA及蛋白表达的影响
RT-PCR结果表明脂氧素A4能抑制LPS刺激的人支气管上皮细胞COX-2 mRNA表达,并呈剂量依赖关系,无脂氧素A4组与对照组相比显著差异(P<0.01),脂氧素A4组与无脂氧素A4组相比差异显著(P<0.01),100 μmol/L与400 μmol/L脂氧素A4组相比差异有统计学意义(P<0.01),见图3。 正常人支气管上皮细胞中COX-2蛋白有微量表达,脂氧素A4抑制LPS刺激的人支气管上皮细胞的COX-2蛋白表达,也呈浓度依赖关系,无脂氧素A4组与对照组相比差异有显著(P<0.01),脂氧素A4组与无脂氧素A4组相比差异有显著(P<0.01),100 μmol/L与400 μmol/L脂氧素A4组相比差异显著(P<0.01),见图 4。
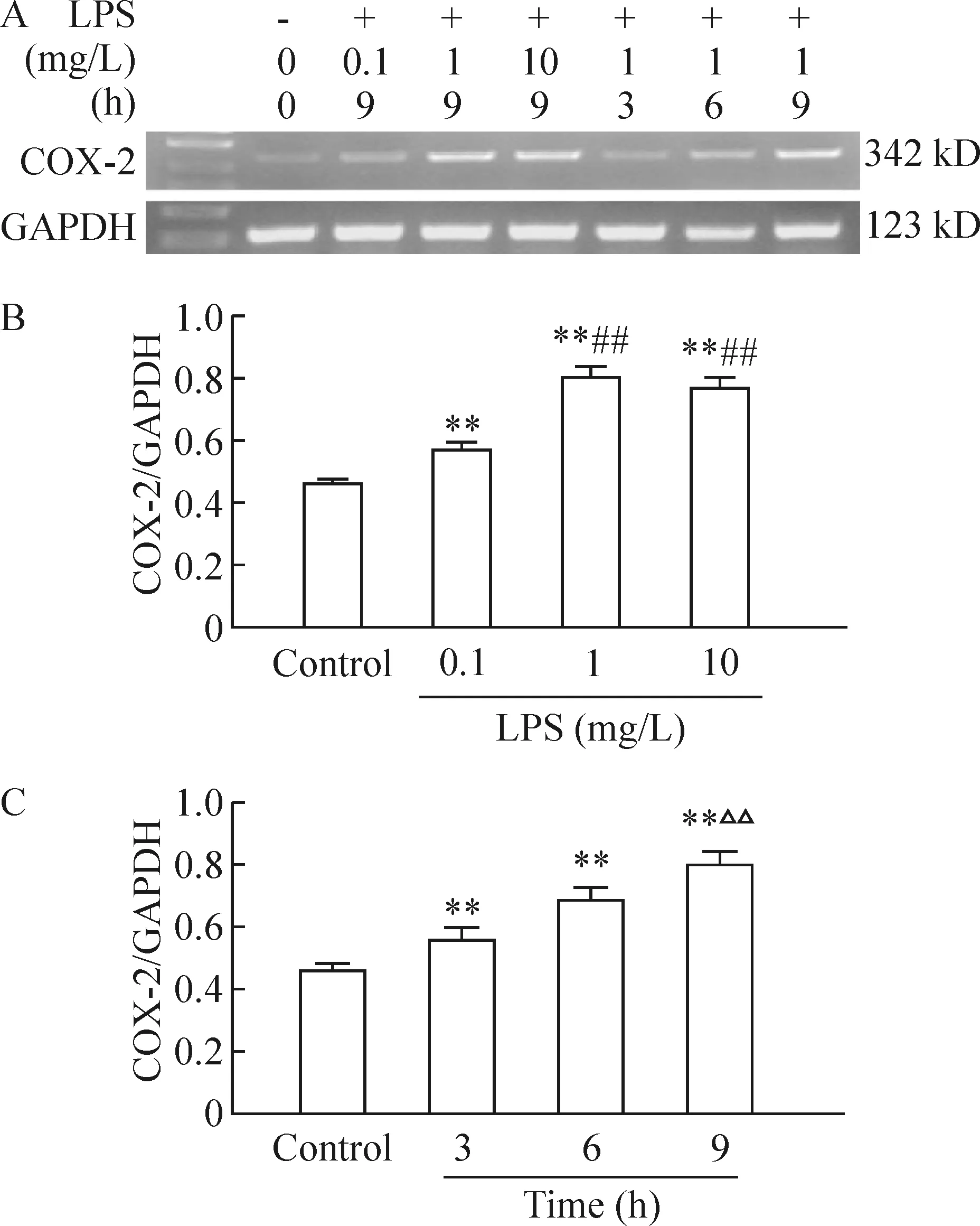
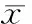
图1HBECs在不同浓度(0.1、1、10mg/L)LPS刺激9h后及在1mg/LLPS刺激不同时间(3h、6h、9h)后COX-2mRNA的表达
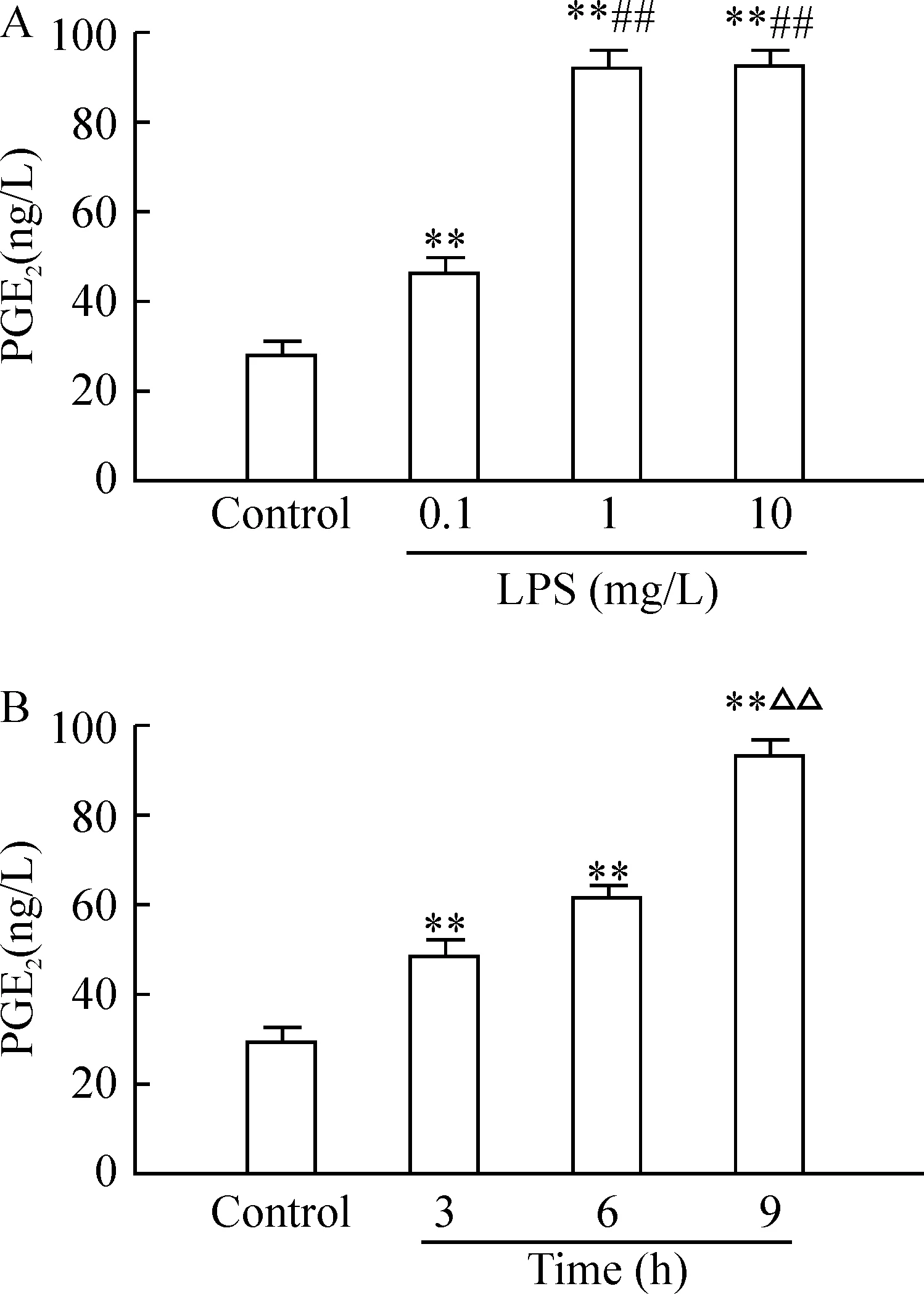

图2HBECs在不同浓度(0.1、1、10mg/L)LPS刺激9h和1mg/LLPS刺激不同时点(3、6、9h)后上清液PGE2的水平
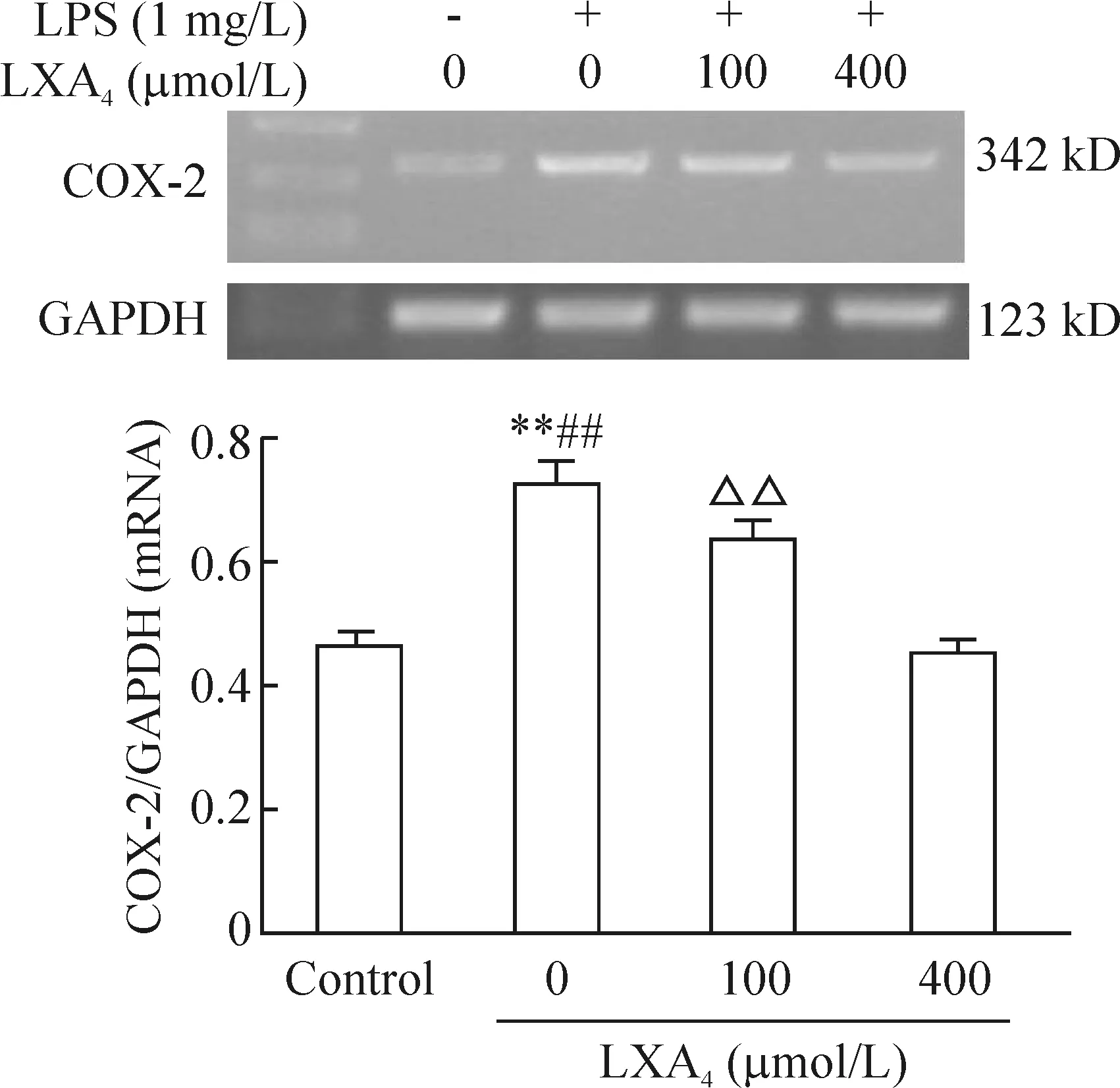
图3脂氧素A4对LPS刺激的人支气管上皮细胞COX-2mRNA的影响
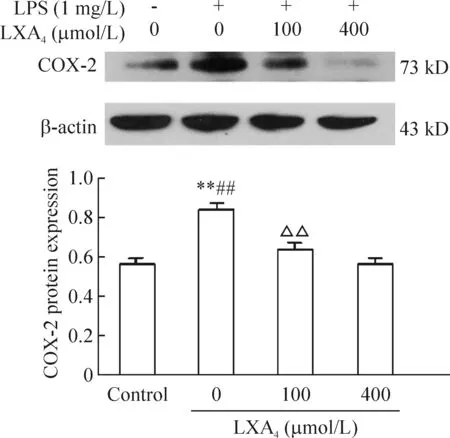
图4脂氧素A4对LPS刺激人支气管上皮细胞COX2蛋白表达的影响
3脂氧素A4对于LPS刺激的人支气管上皮细胞上清液PGE2含量的影响
脂氧素A4能抑制HBECs COX-2表达,PG因此我们推测脂氧素A4是否也能抑制其产物前列腺素E2的表达。HBECs上清 ELISA检测发现,脂氧素A4能抑制LPS刺激后人支气管上皮细胞上清中 PGE2的含量,并呈剂量依赖关系。无脂氧素A4组与对照组相比差异显著(P<0.01),人支气管上皮细胞上清液PGE2从无脂氧素A4组的(97.019±1.425)ng/L降至100 μmol/L脂氧素A4组的(54.077±3.141)ng/L 以及400 μmol/L脂氧素A4组的(39.187±0.775)ng/L (P<0.01)。100 μmol/L与400 μmol/L脂氧素A4组相比差异显著(P<0.01),见图5。
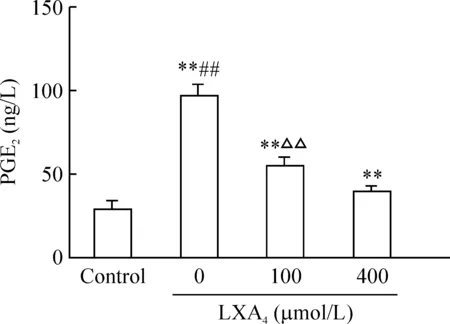
图5不同浓度脂氧素A4对HBECsPGE2表达影响
4相关性分析
Pearson相关分析结果表明脂氧素A4抑制LPS诱导的COX-2蛋白表达与前列腺素E2水平呈相关性(r=0.99,P<0.01)。
讨 论
急性肺损伤及ARDS是临床常见急危重症,病死率极高[7]。目前从针对内毒素性肺损伤的治疗中,我们不难发现抑制炎症启动阶段的促炎介质释放是主旋律[8]。新近研究发现炎症是一个从启动到消退的程序化过程,机体还存在一整套的炎症自限机制来精密调控炎症的发展和消退[9]。在此基础上提出的“促炎症消退”的炎症治疗新策略已成为新的研究热点。
在呼吸系统炎症中前列腺素类物质如PGE2作为主要的炎症介质,起到关键作用[10]。前列腺素类物质属于类花生酸家族,是花生四烯酸经过COX催化而产生[10]。COX是花生四烯酸转化成类花生酸类物质途径中重要的限速酶[4]。机体中主要有2种COX异构体:COX-1及COX-2[11]。COX-1属于管家酶,在大多数组织和细胞中均有表达,其表达相对恒定[12]。 COX-2 在多数情况下处于静息状态或低表达,只有在炎症反应或其它病理条件下才大量表达[13]。在肺部炎症中也存在COX-2 以及PGE2的高表达[4]。
在炎症刺激下,人肺上皮细胞COX-2表达增加[3]。另外,COX-2基因敲除的小鼠炎症反应性增加[14]。由于人支气管上皮细胞表达Toll样受体4并能分泌PGE2[15],因此本研究应用LPS刺激体外培养的人支气管上皮细胞,并且利用MTT法(结果省略)发现1 mg/L LPS刺激细胞可以建立体外炎症模型。目前的研究中,我们也发现人支气管上皮细胞在LPS直接刺激下COX-2 mRNA及PGE2的表达增加(与MTT结果相符),而且我们更进一步的研究结果表明COX-2表达呈时间、剂量依赖关系。
脂氧素是调节炎症消退的重要抗炎介质,在对肺部疾病(如哮喘)体外及体内研究都表明脂氧素具有抗炎作用[16-17]。有研究报道预先给予脂氧素类似物能减轻内毒素所致的小鼠急性肺损伤程度[18]。Medeiros等[19]报道脂氧素A4同样能抑制内毒素所致大鼠眼葡萄膜炎的环氧合酶表达及其活性。另外据报道脂氧素能抑制人肺上皮细胞损伤后的炎症反应并促进其向稳态表型转化[15]。本研究发现脂氧素能抑制LPS刺激培养下人支气管上皮细胞COX-2 mRNA及蛋白的表达,也同时能抑制其上清液PGE2水平,且存在剂量依赖关系。数据相关性分析结果表明脂氧素A4抑制LPS诱导的COX-2表达与PGE2产物具有相关性。因此脂氧素能抑制LPS PGE2产物可能是由于其抑制LPS诱导COX-2表达所致。综上所述我们认为脂氧素A4可能部分通过依赖COX-2的方式促进肺部炎症消退,为急性肺损伤的治疗提供一个新的治疗靶点。
[1] Udobi KF, Childs E, Touijer K. Acute respiratory distress syndrome[J]. Am Fam Physician, 2003, 67(2): 315-322.
[2] Martin LD, Rochelle LG, Fischer BM, et al. Airway epithelium as an effector of inflammation:molecular regulation of secondary mediators[J]. Eur Respir J, 1997,10(9): 2139-2146.
[3] Bonnans C, Fukunaga K, Levy MA, et al. Lipoxin A4regulates bronchial epithelial cell responses to acid injury[J]. Am J Pathol, 2006, 168(4):1064-1072.
[4] Ricciotti E, Fitzgerald GA. Prostaglandins and inflammation[J]. Arterioscler Thromb Vasc Biol, 2011, 31(5):986-1000.
[5] Kühn H, O’Donnell VB. Inflammation and immune regulation by 12/15-lipoxygenases[J]. Prog Lipid Res, 2006, 45(4):334-356.
[6] Romano M. Lipoxin and aspirin-triggered lipoxins[J]. Scientific World J, 2010, 10: 1048-1064.
[7] Sen S, Sen S, Sentürk E, et al. Postresectional lung injury in thoracic surgery pre and intraoperative risk factors: a retrospective clinical study of a hundred forty-three cases[J]. J Cardiothorac Surg, 2010, 5:62.
[8] Scher JU, Pillinger MH. The anti-inflammatory effects of prostaglandins[J]. J Investig Med, 2009, 57(6):703-708.
[9] Spite M, Serhan CN. Novel lipid mediators promote resolution of acute inflammation impact of aspirin and statins[J]. Circ Res, 2010, 107(10):1170-1184.
[10]Park JY, Pillinger MH, Abramson SB. Prostaglandin E2synthesis and secretion: the role of PGE2synthases[J]. Clin Immunol, 2006, 119(3):229-240.
[11]Morita I. Distinct functions of COX-1 and COX-2[J]. Prostaglandins Other Lipid Mediat, 2002, 68-69:165-175.
[12]Turini ME, DuBois RN. Cyclooxygenase-2: a therapeutic target[J]. Annu Rev Med, 2002, 53:35-57.
[13]Fortier MA, Krishnaswamy K, Danyod G, et al. A postgenomic integrated view of prostaglandins in reproduction:implications for other body systems[J]. J Physiol Pharmacol, 2008, 59(Suppl 1):65-89.
[14]Bonner JC, Rice AB, Ingram JL, et al. Susceptibility of cyclooxygenase-2-deficient mice to pulmonary fibrogenesis[J]. Am J Pathol, 2002, 161(2):459-470.
[15]Bonnans C, Gras D, Chavis C, et al. Synthesis and anti-inflammatory effect of lipoxins in human airway epithelial cells[J]. Biomed Pharmacother, 2007, 61(5):261-267.
[16]Levy BD, De Sanctis GT, Devchand PR, et al. Multi-pronged inhibition of airway hyper-responsiveness and inflammation by lipoxin A4[J]. Nat Med, 2002, 8(9):1018-1023.
[17]Planaguma A,Kazani1 S, Marigowda1 G, et al. Airway lipoxin A4generation and lipoxin A4receptor expression are decreased in severe asthma[J]. Am J Respir Crit Care Med, 2008,178(6): 574-582.
[18]Jin SW, Zhang L, Lian QQ, et al. Posttreatment with aspirin-triggered lipoxin A4analog attenuates lipopolysaccharide-induced acute lung injury in mice: the role of heme oxygenase-1[J]. Anesth Analg, 2007, 104(2):369-377.
[19]Medeiros R, Rodrigues GB, Figueiredo CP, et al. Molecular mechanisms of topical anti-inflammatory effects of lipoxin A4in endotoxin-induced uveitis[J]. Mol Pharmacol, 2008, 74(1): 154-161.
LipoxinA4reduceslipopolysaccharide-inducedexpressionofcyclooxygenase2andprostaglandinE2inhumanbronchialepithelialcells
ZHU Tian-qi1, ZHENG Sheng-xing2, JIN Sheng-wei3
(1DepartmentofAnesthesia,TheFirstAffiliatedHospitalofWenzhouMedicalCollege,2DepartmentofAnesthesia,3IntensiveCareUnit,TheSecondAffiliatedHospitalofWenzhouMedicalCollege,Wenzhou325000,China.E-mail:jinshengwei69@yahoo.com.cn)
AIM: To explore the effects of lipoxin A4on the expression of cyclooxygenase 2 (COX-2) in human bronchial epithelial cells (HBECs).METHODSHBECs were incubated with various concentrations (0.1, 1 and 10 mg/L) of lipopolysaccharide(LPS) for 9 h, or 1 mg/L LPS for different time (3 h, 6 h and 9 h). The levels of COX-2 mRNA in HBECs and prostaglandin E2(PGE2) in the culture supernatant were measured. In addition, the HBECs were exposed to lipoxin A4at concentration of 0, 100 and 400 μmol/L after stimulated with LPS at concentration of 1 mg/L for 9 h, and the supernatant of the culture cells was collected for determining the content of PGE2by ELISA. The cells were also harvested, and the mRNA and protein levels of COX-2 were analyzed by RT-PCR and Western blotting, respectively.RESULTSLPS increased the mRNA expression of COX-2 and production of PGE2in a dose and time dependent manners in HBECs. Induction of COX-2 mRNA and protein by LPS were inhibited by lipoxin A4in a dose-dependent manner. Lipoxin A4also significantly decreased LPS-induced production of PGE2.CONCLUSIONLipoxin A4down-regulates LPS-induced expression of COX-2 and consequently inhibits the production of PGE2in HBECs.
Lipoxin A4; Human bronchial epithelial cells; Cyclooxygenase 2; Lipopolysaccharides
R332.2
A
10.3969/j.issn.1000-4718.2012.03.016
猜你喜欢
保健医苑(2022年1期)2022-08-30
中老年保健(2021年11期)2021-08-22
现代临床医学(2021年4期)2021-07-31
昆明医科大学学报(2021年3期)2021-07-22
猪业科学(2021年3期)2021-05-21
医学前沿(2021年18期)2021-04-14
心肺血管病杂志(2020年5期)2021-01-14
家庭医药(2020年12期)2020-12-28
家庭医药(2020年23期)2020-01-03
家庭医学(2018年1期)2018-03-09
