
注意资源数量对内隐序列学习的影响:来自ERP的证据*
2012-10-17 07:18刘静远钟毅平
湖北科技学院学报 2012年5期
刘静远,钟毅平
(湖南师范大学教育科学学院,湖南长沙410081)
注意资源数量对内隐序列学习的影响:来自ERP的证据*
刘静远,钟毅平
(湖南师范大学教育科学学院,湖南长沙410081)
内隐学习有别于传统的外显学习,它是一种不知不觉的学习过程,是有机体与环境接触时,无目的、自动地获得事件或客体间结构关系的过程。内隐序列学习是内隐学习研究的主要范式之一。目前关于序列学习的脑机制的研究还比较薄弱,利用脑电事件相关电位(ERP)对序列学习进行的研究还比较少。本研究运用ERP的技术手段,通过操纵分心刺激的数量来考察注意资源数量对内隐序列学习的影响,结果从电生理层面支持了Shanks和Channo(2002)的观点。
注意资源;内隐;序列学习;ERP
一、研究背景
1.内隐序列学习
内隐学习有别于传统的外显学习,它是一种不知不觉的学习过程,是有机体与环境接触时,无目的、自动地获得事件或客体间结构关系的过程[1]。第一篇以内隐学习为题的论文,是美国心理学家A.S.Reber于1967年发表的文章“人工语法的内隐学习”(Implicit Learning of Artificial Grammar),在这篇文章中,内隐学习的概念被首次提出,内隐学习由此正式成为心理学研究者们关注的对象[2]。
有序地产生有组织的行为是人类心理机能的基本特点。获得关于事件或行为的序列知识对于人们的日常学习和生活至关重要[3]。近年来,有关内隐学习的研究发现,人们在序列学习中可以获得无意识知识,这些知识虽然会影响行为,但是人们却意识不到它们[4~6]。大量实验表明,序列学习的习得行为表现可以独立于外显知识的获得,即:即使被试并未意识到序列规则的存在,也不能对其正确再现,其行为仍可体现出学习效应(反应时的获益)[7~9]。内隐序列学习作为研究内隐学习的一个主要范式,是有其优势的,比如,被试并不知道序列存在规律,因此对它的知识获得与提取过程都是内隐的,与人工语法相比,存在着较高的一致性与连贯性。所以,序列学习一直是内隐学习的一个热点[10]。
2.抑制表达理论
目前在内隐序列学习的研究中,有关序列学习获得何种知识以及注意在序列学习中的作用,研究者之间还存在很大的争议[3,9,11-15]。在以往研究的基础上Frensch指出,尽管许多有关内隐序列学习的研究都表明,被试在单任务条件下的学习成绩明显好于双任务条件下的学习成绩,但是即使在双任务条件下内隐学习效应也并没有完全消失而只是略小于单任务条件[16]。由此,他们提出,应进一步区分第二任务对序列学习本身和学习结果表达的影响,如果第二任务既影响序列学习本身又影响学习结果的表达,那么在不同的学习条件下被试的学习成绩应有所不同。然而,Frensch、Lin和Buchner对此进行的研究结果却表明,当三组被试在学习阶段分别完成数量不同的任务组段时,无论在“单任务”条件还是“双任务”条件下,他们的学习成绩都无显著差异[17]。因而,研究者认为,第二任务只会抑制序列学习的内隐序列学习与注意的关系表达,但并不会影响序列学习本身。
Heuer和Schmidtke的研究为抑制表达理论提供了初步的支持,他们的结果表明,在单任务测试条件下,接受单任务学习条件的被试的学习成绩高于接受双任务学习条件的被试[18]。此外,Schvaneveldt和Gomez利用或然序列进行的研究,也为Frensch等人的理论提供了进一步的支持[19]。
不过,Frensch等人的研究受到了Shanks和Channon的质疑,Shanks和Channon对Frensch等人的实验设计进行了改进[20],结果发现,当以数声任务作为第二任务时,无论被试在“单任务”还是“双任务”条件下进行学习或测试,被试的序列学习成绩都会显著受到第二任务影响。因此,他们认为,内隐序列学习需要注意资源的参与。Shanks(2003)的研究结果也为这一观点提供了进一步的支持[11]。
3.问题的提出
这些研究者在探讨注意的影响时,主要都是采用的双任务范式,通过增加的次级任务来分析内隐序列学习是否需要注意资源。而本研究试图从操控序列本身的角度出发,通过操纵目标刺激周围的分心刺激的数量来考察注意资源数量对目标刺激的影响,并通过ERP的技术手段,尝试从电生理的层面来检验抑制表达理论,探讨注意资源对内隐序列学习的影响。
本研究选取的ERP指标:
ERN
ERN(error-related negativity)是额骨中心区在一个不正确(不符合预期)的反应过后约100ms时发生的否定性反应[21]。ERN通常被作为检测到错误[22]或者反应发生冲突的指标[23]。序列学习过程中,被试除了对呈现的刺激进行预期外,很可能同时进行纠错,这就用到了ERN。
P300
P3是顶骨的颅侧区在刺激过后约300-400ms时发生的显性反应,通常与刺激违反了期望的程度相关[24,25]。在序列学习研究中,研究者通常在规则序列中插入偏差刺激,这种偏差刺激常常会引发P300的变化。
根据Shanks和Channon对“抑制表达”理论的质疑[20],由于注意资源的被占用,内隐序列学习效果会受到分心刺激数量的影响。随着分心刺激数量的增加,被试的内隐学习也会出现削弱。具体而言,由于P300和ERN都是异常刺激的指标(异常刺激相比标准刺激,会引起更大的P300和ERN),如果内隐序列学习需要较多的注意资源,可以推知,分心刺激占用的注意资源越多,则投入到内隐序列学习中的注意资源就越少,那么在完成任务过程中投入注意资源最多的组在出现异常刺激时,由于对内隐序列的习得较差,对异常刺激的敏感度较小,就会出现较小的P300和ERN波幅。
二、实验
1.目的
通过操纵分心刺激的数量,考察内隐序列学习效果是否会受分心刺激数量的影响。
2.设计
单因素(分心刺激数量:无分心刺激vs.单分心刺激vs.双分心刺激)被试内设计。
3.假设
无分心刺激组(A组)在出现异常刺激时会出现较大的P300和ERN波幅;双分心刺激组(C组)在出现异常刺激时会出现较小的P300和ERN波幅;单分心刺激组(B组)在出现异常刺激时出现的P300和ERN波幅介于A、C组之间。
4.被试
湖南师范大学19名本科生,10名女性,年龄在18~23岁之间,平均年龄19岁3个月。被试均为右利手,身体健康无神经系统疾病,没有脑部损伤史,视力正常或矫正后正常。自愿参加本实验,母语皆为汉语。实验完成后付给一定报酬。其中一名女性被试由于实验时不认真(正确率仅56.9%)数据被剔除。
5.刺激材料与环境
刺激物为圆形。圆的直径为2cm,由分辨率为1280×1024电脑屏幕呈现,屏幕距被试双眼距离80cm。被试坐在舒适的座椅上,并避免除按键反应外的身体动作。
以下三个图片分别为无分心刺激物示意图、单分心刺激物示意图、双分心刺激物示意图。

图1 刺激物示意图
(实验中,被试的任务是:发现在哪条线段的上方出现了圆,就对这条线段进行指认;因此,出现在线段下方的圆与任务是无关的,属于分心刺激。)
6.实验流程
实验阶段,被试根据屏幕上刺激出现的位置按键盘上的相应键,从左到右四个圆点分别对应于键盘上的D、F、J、K键(如示意图)。刺激物按以下顺序呈现(1—4代表从左到右的四个圆点):SOC1=4-1-3-4-2-3-2-1-4-3-1-2-(4)-(1)-(3)。该序列为二阶序列(second—order conditional quenees,SOC)。二阶序列中,连续两个刺激位置决定下一个刺激的位置,所以在此SOC1序列中,前两个刺激依次出现在4,1位置,则下一个刺激一定出现在3,依此类推。连续三个位置构成的子序列称为序列组块(如4-1-3为一个序列组块,1-3-4为另一个序列组块),SOC序列中有多少个数,就有多少个不同的组块。因此在本研究所用的序列中,存在4-1-3,1-3-4,3-4-2,4-2-3,2-3-2,3-2-1,2-1-4,1-4-3,4-3-1,3-1-2,1-2-4,2-4-1这12个固定组合,出现前两个数,则确定了第三个数。
SOC序列可以从任何一个位置开始,循环进行,其中每个位置出现的概率均等。实验中,90%的刺激为标准刺激,标准刺激出现符合SOC1的序列规则。另有10%的刺激为异常刺激,异常刺激来自以下序列:SOC2:4-1-3-4-2-3-2-1-4-3-1-2-(4)-(1)-(2)。异常刺激出现的位置是随机的。对每个组块(如前两个刺激为4-1),都有90%的刺激为标准刺激(3—4—2),10%为异常刺激(3—4—1)。
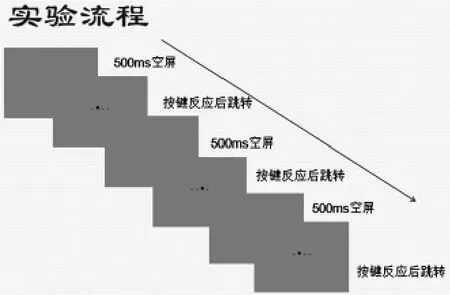
图2 实验流程图
实验中,被试者并不知道刺激遵循一定规律,而仅被要求尽量既准又快地按键。反应任务包括9个组段(block),每个组段有120个刺激(trial),其中第一、二个刺激的位置是随机的,随后将按SOC1序列规律(10%为异常)进行。反应刺激间隔为500ms,每个组段之间休息30s。刺激物出现在屏幕四个位置上,从左到右四个位置分别对应键盘上的D、F、J、K键(如图中圆点出现在从左数第二个位置,应按F键进行反应)。9个组段的呈现顺序为(A-B-C-A-B-C-A-B-C),对各组的呈现顺序进行平衡。
任务完成后,对被试进行调查,检测其是否已知晓实验目的。
7.EEG记录
在双眼外侧安置电极记录水平眼电(HEOG),左眼上下安置电极记录垂直眼电(VEOG)。每个电极处的头皮电阻保持在5 kΩ以下。采样频率为256Hz。数据经离线处理后以所有导联的平均电位作为参考,并通过0.05~35Hz离线滤波。
8.行为数据分析
使用spss18.0对被试的正确反应的反应时进行单因素(标准刺激vs异常刺激)重复测量方差分析。分析中,排除反应时小于等于200ms或大于等于1000ms的trial,以及按键反应错误的trial。
9.ERP数据分析
刺激出现前100ms到刺激出现后700ms为一个时程。以刺激前100ms到刺激出现的平均幅度作为基线。将全部导连的数据的平均值作为参考,每导连的响应由该导连的原始信号减去参考得到。将所有被试(无分心刺激/单分心刺激/双分心刺激组)对标准、异常刺激的脑电响应分别进行总平均,得到(无分心刺激/单分心刺激/双分心刺激)各组标准与异常刺激的ERP波形。
垂直或水平眼点幅度超过80 μV的刺激、反应时小于等于200ms或大于1000ms的刺激,以及按键反应错误的刺激均被剔除。被剔除的刺激数控制在10%以内。比较无分心刺激/单分心刺激/双分心刺激组块的ERP波形,对ERN幅度(150~250ms时间窗内的平均电位)以及P300幅度(300~600ms时间窗内的平均电位)进行3(学习类型:无分心刺激/单分心刺激/双分心刺激)×2(刺激类型:标准/异常)×3(导联:Fz/Cz/Pz)方差分析。
10.结果
实验结束后,询问被试情况,被试均表示在实验过程中不知道实验的目的,并且在外显层面上没有报告出二阶序列规律。
正确率
当出现异常刺激时,各组正确率:A组97.2%,B组97.9%,C组98.6%,三者间差异均不显著;正常刺激的正确率为98.2%,与错误刺激的正确率相比差异不显著。
反应时
正常刺激平均反应时:488.6110ms,异常刺激平均反应时486.6962ms,无显著差异。
A组无分心刺激条件下对异常刺激的平均反应时为468.1439ms,B组单分心刺激条件下对异常刺激的平均反应时为502.6978ms,C组双分心刺激条件下对异常刺激的平均反应时为490.4681ms,组间差异显著F(2,16)=5.96,p<0.05。
ERP波形

图3 标准刺激vs.异常刺激波形图
叠加过后的标准刺激的ERN波幅峰值为-2.021μV,异常刺激的ERN波幅峰值为-2.401μV,两者差异显著F(1,17)=8.41,p<0.01。
标准刺激的P3波幅峰值为5.262μV,异常刺激的P3波幅峰值为5.630μV,两者差异显著F(1,17)=5.46,p<0.05。

图4 刺激物波形图
无分心刺激组的ERN波幅峰值为-5.218μV,单分心刺激组的ERN波幅峰值为-4.493μV,双分心刺激组的ERN波幅峰值为-3.286μV,三组中各组间均差异显著F(2,16)=4.46,p<0.05。
无分心刺激组的P3波幅峰值为8.743μV,单分心刺激组的ERN波幅峰值为7.636μV,双分心刺激组的ERN波幅峰值为7.312μV。A组与B组差异显著F(1,11)=9.66,p<0.01;A组与C组差异显著F(1,11)=9.76,p<0.01;B组与C组间差异不显著。
11.讨论
从结果上看,三组的正确率之间差异不显著,但是可以看到微小的差异,即A组正确率97.2%<B组正确率97.9%<C组正确率98.6%,这可能是天花板效应造成的(即各组正确率均较高),也可能只是被试对三组刺激的反应在外显(反应时)层面上没有区别。标准刺激与异常刺激的正确率之间没有显著差异也可能是这些原因造成的。
再来看各组对异常刺激的反应时上的差异,A组比B组、C组的反应时都要短,这说明A组在任务过程中受到的干扰是最小的,B组和C组的分心刺激确实对被试的外显表现产生了干扰作用。
在电生理的层面上,标准刺激的ERN波幅和P3波幅均显著小于异常刺激的ERN波幅和P3波幅,这说明被试检测出了预设的“异常刺激”,而由于被试在外显层面上没有习得序列规则,说明被试的这种对“异常刺激”的检测是建立在内隐的序列学习的基础上的。
从ERN波幅上来看,A组ERN波幅>B组ERN波幅>C组ERN波幅,相当于对异常刺激的检测能力有:A组>B组>C组,这说明由于注意资源的被占用,内隐序列学习的效果了受到分心刺激数量的影响,即分心刺激数量的增多会导致内隐序列学习的效果变差。P3波幅的对比说明内隐序列学习的效果受到了分心刺激的影响。
三、结论
本实验的假设是根据Shanks和Channon对“抑制表达”理论的质疑[22]而提出的,实验结果表明,在ERN波幅上,由于注意资源的被占用,内隐序列学习效果会受到分心刺激的数量的影响。随着分心刺激数量的增多,被试对目标刺激的内隐学习效果也会出现削弱。具体而言,由于分心刺激占用的注意资源A组<B组<C组,因此在投入到内隐序列学习中的注意资源就有A组>B组>C组,因此A组在出现异常刺激时就会出现较大的ERN波幅(内隐序列习得较快,对异常刺激的敏感度较大)。如此,本实验的结果在电生理层面上支持了Shanks和Channon的观点[20]。
[1]郭秀艳.内隐学习:一种不知不觉的学习[J].教育科学,2003,19(6):41~44.
[2]Reber,A.S.Implicit learning of art ificial grammars[J].Journal of Verbal Learning&Verbal Behavior,1967,(77):317~327.
[3]付秋芳,傅小兰.第二任务对内隐序列学习的影响[J].心理科学,2010,33(4):861~864.
[4]Cleeremans A,Destrebecqz A.,Boyer M.Implicit Learning:News from the front.Trends in Cognitive Science[J].1998,2(10):406~416.
[5]Jiménez L,Méndez C,Cleeremans A.Comparing direct and indirect measures of sequence learning[J].Journal of Experimental Psychology:Learning,Memory,and Cognition,1996,22(4):948~969.
[6]Pacton S,Perruchet P,Fayol M,Cleeremans A.Implicit learning out of the lab:The case of orthographic regularities[J].Journal of Experimental Psychology:Gnenral,2001,130(3):401~426.
[7]Hoffman J,Sebald A,St?cker C.Irrelevant response effects improve serial learning in serial reaction time tasks[J].Journal of Experimental Psychology:Learning,Memory,and Cognition,2001,27(2):470~482.
[8]Jiménez L.Intention,attention,and consciousness in probabilistic sequence learning[M].In:Jiménez L.Attention and implicit learning.Amsterdam:John Benjamins Publishing Company,2003.43~68.
[9]Willingham D B,Wells L A,Farrell J M.Implicit motor sequence learning is represented in response locations[J].Memory&Cognition,2000,(28):366~375.
[10]许昭.内隐序列学习研究方法简评[J].辽宁教育行政学院学报,2008,25(11):57~58.
[11]Shanks DR.Attention and awareness in“implicit”sequence learning.In L.Jiménez.Attention and implicit learning Amsterdam/Philadelphia:John Benjamins,2003.11~42.
[12]葛操,白学军,沈德立.注意负荷对内隐序列学习的影响研究[J].心理科学,2007,30(5):1 029~1 032.
[13]郭秀艳.内隐学习[M].上海:华东师范大学出版社,2003.
[14]付秋芳,傅小兰.内隐序列学习与注意的关系[J].心理科学进展,2006,14(6):817~821.
[15]Deroost N,Coomans D,Soetens E.Perceptual load improves the Expression but not learning of relevant sequence information[J].Experimental Psychology,2009,56(2):84~91.
[16]Frensch P A.One concept,multiple meanings:On how to define the concept of implicit learning.In:Stadler M A,Frensch P A.Handbook of implicit learning[J].Thousand Oaks,CA:Sage,1998,47~104.
[17]Frensch P A,Lin J,Buchner A.Learning versus behavioral expression of the learned:The effects of a secondary tone-counting task on implicit learning in the serial reaction time task[J].Psychological Research,1998,61(1):83~98.
[18]]Heuer H,Schmidke V.Secondary-task effects on sequence learning[J].Psychological Research,1996,59(2):119~133.
[19]Schvaneveldt R W,Gomez R L.Attention and probabilistic sequence learning[J].Psychological Research,1998,61(2):175~190.
[20]Shanks D R,Channon S.Effects of a secondary task on implicit sequence learning:Learning or performance[J].Psychological Research,2002,66(1):99~109.
[21]Gehring W.J.,Goss B.,Coles M.G.,Meyer D.E.,Donchin E.A neural system for error detection and compensation[J].Psychological Science,1993,4(6):385~390.
[22]Scheffers M.K.,Coles M.G.H.,Bernstein P.,Gehring W.J.,Donchin,E.Event-related brain potentials and error-related processing:An analysis of incorrect responses to go and no-go stimuli[J].Psychophysiology,1996,33(1):42~53.
[23]Gehring W.J.,Fencsik D.E.Functions of the medial frontal cortex in the processing of conflict and errors[J].Journal of Neuroscience,2001,21(23):9 430~9 437.
[24]Courchesne E.,Hillyard S.A.,Galambos R.Stimulus novelty,task relevance and the visual evoked potential in man[J].Electroencephalography and Clinical Neurophysiology,1975,39(2):131~143.
[25]Donchin,E.,Coles M.G.Is the P300 component a manifestation of context updating?Behavioral and Brain Sciences,1988,11(3):357~427.
G44
A
1006-5342(2012)05-0037-04
2012-02-20
猜你喜欢
癫痫与神经电生理学杂志(2022年6期)2022-02-10
中华养生保健(2020年7期)2020-11-16
价值工程(2018年25期)2018-09-26
大科技·百科新说(2017年4期)2017-05-13
家教世界·创新阅读(2016年11期)2016-12-27
天津护理(2016年3期)2016-12-01
故事会(2016年15期)2016-08-23
西南交通大学学报(2016年4期)2016-06-15
华北电力大学学报(自然科学版)(2016年3期)2016-04-25
中国工作犬业(2010年11期)2010-11-06
