
白菜与甘蓝之间体细胞杂交种获得与遗传特性鉴定
2012-10-11 02:12廉玉姬
生物工程学报 2012年9期
廉玉姬
临沂大学 生命科学学院,山东 临沂 276005
农业生物技术
白菜与甘蓝之间体细胞杂交种获得与遗传特性鉴定
廉玉姬
临沂大学 生命科学学院,山东 临沂 276005
为拓宽白菜育种的基因资源,改良白菜品质,以白菜 (Brassica campestris,2n=20,AA) 和甘蓝(B. oleraceaL. var.capitata,2n=18,CC) 的子叶和下胚轴为材料分离、制备原生质体。采用 40%聚乙二醇(Polyethylene glycol,PEG) 进行原生质体融合。融合细胞在以0.3 mol/L蔗糖、0.3 mol/L葡萄糖为渗透稳定剂,附加1.0 mg/L 2,4-D + 0.5 mg/L 6-苄氨基嘌呤 (6-BA) + 0.1 mg/L 1-萘乙酸 (NAA) + 1.0 mg/L激动素 (Kinetin,Kin) 的改良K8p培养基中培养并诱导细胞分裂。小愈伤组织经增殖培养后在MS + 0.2 mg/L玉米素 (Zeatin,ZEA) + 1 mg/L 6-BA + 0.5 mg/L Kin + 0.4 mg/L NAA的固体分化培养基上诱导出不定芽。30 d后再转入MS基本培养基, 获得完整的再生植株。将生根的植株转移到花盆,并对其杂种性质进行形态学、细胞学和分子生物学鉴定。结果表明,经细胞融合分裂出的320个愈伤组织中,获得了35棵再生植株,其再生率达10.94%。形态学观察显示,绝大多数再生植株的叶面积较大,株型和叶型为两种杂交亲本的中间型,部分植株的叶片浓绿、肥厚。染色体计数结果显示,36.4%的再生植株染色体数为2n=38;36.4%的再生植株的染色体数为2n=58~60;27.2%的再生植株的染色体数为2n=70~76, 超过两个融合亲本的染色体数的总和。流式细胞仪测定DNA 含量显示,再生植株 DNA 含量变化比较大,其结果与染色体鉴定结果相吻合。随机扩增多态性 DNA (Random amplified polymorphic DNA,RAPD) 和基因组原位杂交 (Genomicin situhybridization,GISH) 分析结果证明再生植株具有双亲基因组。体细胞杂种花粉育性比较低,杂交、回交后其育性逐渐获得恢复,与白菜回交后代逐渐恢复了育性。通过体细胞杂交和回交、杂交获得了形态变化广泛的个体,为白菜的品种育种提供多样的种质资源。
体细胞杂交,植株再生,基因组原位杂交,随机扩增多态性DNA,染色体数,后代
Abstract:In order to broaden Chinese cabbage gene pool, we conducted interspecific somatic hybridization between Chinese cabbage (Brassica campestris, 2n=20, AA) and Cabbage (B. oleracea,2n=18, CC).Protoplasts were isolated from 10-day-old cotyledons and hypocotyls of young seedlings, and fused by 40% polyethylene glycol (PEG). Fused cells were cultured in modified K8p liquid medium supplemented with some plant growth regulators. Fusion products were characterized by their morphological, cytological and molecular biological traits. The results showed that, a total of 35 regenerated green plants were obtained from 320 calli, the plant regeneration frequency was 10.94%, and eleven of which were survived in greenhouse. All regenerants were true hybrids as confirmed by randomly amplified polymorphic DNA (RAPD) and genomicin situhybridization(GISH) analysis. Ploidy levels of hybrid plants were determined by chromosome counting and flow cytometry. The sum of the chromosome number (2n= 38) from the two fusion patents were found in 36.4% of regeneratns; another 36.4% had chromosomes range to 58-60; 27.2% had more chromosomes ranges to 70-76. All regenerated plants produced normal flowers. We investigated the pollen fertility and seed set after self-pollination and backcrossing with the parental species. For hybrids with chromosomes more than 38 it was possible to obtain some seeds when they after self-pollination. Within the group of hybrids with 38 chromosomes, seed set were very variable, only 0.11 seeds per pod by self-pollination, 0.23-0.76 by open-pollination, 0.02-0.04 by backcrossing with Chinese cabbage. Progeny lines obtained by self-pollination had larger leaves and leaf shapes intermediate of the parental species. Pollen fertility was gradually recovered in the first and second progenies. The backcrossing progeny lines,as a whole, exhibited morphologies were similar to Chinese cabbage. Morphological variations were observed among the somatic hybrids and their progenies.
Keywords:somatic hybridization, regeneration, genomicin situhybridization, random amplified polymorphic DNA,chromosome number, progeny
Introduction
The enrichment of cultivated gene pools via introgression of favorable genes/gene complexes from wild allies is currently a priority in many crop improvement programs. Extensive genomic resources in the Brassicaceae family, to which the genusBrassicabelongs, provide the basis for this improvement. However, the incompatibility between most wild and cultivated species, coupled with low fertility in F1 hybrids, severely restrain the chances of introgression of desirable traits[1-2]. Somatic hybridization provides a means to overcome sexual incompatibility and has been used to obtain many intraspecific, interspecific, intergeneric, intertribal,and even interfamilial somatic hybrids[3-6]. This technology allows intrageneric hybridizations as well as the production of intergeneric hybrids and cybrids[7].
Protoplast technology has been extensively applied in theBrassicaspecies by fusing protoplasts fromB. campestrisandB. oleraceato widen their genetic diversity, among other things[8-10]. Other examples of interspecific somatic hybridization betweenB. campestrisandB. oleraceahave been reported, including the transfer of economic traits such as cytoplasmic male sterility[11-14]and disease resistance[15-16]. In this present study, somatic hybrids that have valuable traits from both Chinese cabbage and cabbage were producedto broaden theirgene pool. The somatic hybrids and their progenies were characterized by investigating their morphological, cytological, and molecular biological characteristics using flow cytometry, random amplified polymorphic DNA (RAPD), genomicin situhybridization (GISH), and plant morphological comparison.
1 Materials and methods
1.1 Plant materials
Inbred lines ofB. oleraceaL. var.italica(cabbage), andB. campestris(Chinese cabbage)were used as plant materials in somatic hybridization. The seeds were surface-sterilized using 70% ethyl alcohol for 30s, followed by 15 min in 50% commercial Chlorox bleach solution.Exactly two drops of Tween-20 were added to the seeds, which were then rinsed three times with sterile distilled water. The sterilized seeds were germinated and propagatedin vitroon a Murashige and Skoog (MS)[17]medium supplemented with 3%sucrose and solidified with 0.8% agar under controlled conditions (25 °C, 16 h photoperiod,84 μmol/(m2·s), and under white fluorescent light).
1.2 Protoplast isolation, fusion, culture, and plant regeneration
Protoplasts of Chinese cabbage and cabbage were isolated from 10-day-old seedlings using an enzyme solution containing 0.4 mol/L mannitol,50 mmol/L CaCl2×2H2O, 1% cellulysin (Calbiochem,USA), and 0.5% macerozyme (Calbiochem, USA) at pH 5.8. Protoplast isolation and fusion were performed as described by Lian and Lim[18]. The fused protoplasts were cultured in modified K8p medium[19]supplemented with 1 mg/L 2,4-dichorophenoxyacetic acid (2,4-D), 0.5 mg/L 6-benzylaminopurine (6-BA), 0.1 mg/L 1-naphthaleneacetic acid (NAA), and 1 mg/L Kinetin (Kin) to promote cell division. The protoplasts were first cultured in 6 mm plastic Petri dishes containing 1 mL to 1.5 mL liquid culture medium. The Petri dishes were then sealed with Parafilm®and incubated at 25 °C in the dark. When the small calli reached diameters of 2 mm to 3 mm, they were transferred to the regeneration medium containing 0.2 mg/L Zeatin(ZEA), 1 mg/L 6-BA, 0.5 mg/L Kin, and 0.4 mg/L NAA. The calli were solidified by adding 8 g/L agar at pH 5.8 for shoot regeneration at 25 °C under fluorescent light at 84 μmol/(m2·s) and a 16 h photoperiod. The calli were transferred to a new medium every 2 to 3 weeks, and the regenerated shoots were transferred to an MS basal medium supplemented with 0.2 mg/L NAA for growth and rooting.
1.3 Ploidy estimation using flow cytometry
The fluorescence of the samples was measured on a Partec flow cytometer (Partec PA-I, Germany)equipped with a high-pressure mercury lamp. A total of 0.2 g of fresh leaves from the regenerated plants and fusion parents were excised, chopped, and incubated in 2 mL of the nuclear extraction buffer(high-resolution DNA kit type P, Solution A; Partec)for 1 min. The resulting mixture was then filtered for 30 min using Partec CelltricsTMand then stained for 2 min with 1 mL of Partec HR-B solution. TheB. oleraceaandB. campestrisdiploids were used as controls, against which the relative fluorescence intensities from the regenerated plants were compared.
1.4 Random amplified polymorphic DNA (RAPD)analysis
Total DNA was isolated from the leaves of greenhouse-grown parental lines and 11 regenerated plants following the protocols of the cetyltrimethylammonium bromide (CTAB)method[20]. RAPD analysis was also conducted on 10 putative somatic hybrids and protoplast fusion parents. In total, 30 primers (Operon Technologies,USA) were tested to identify those that can produce specific bands in both fusion parents. The amplification conditions were 35 cycles of 94 °C for 40s, 40 °C for 60s, and 72 °C for 60s.Amplification products were analyzed by electrophoresis in 1.0% (W/V) agarose gel and detected by staining with ethidium bromide. Thegels were photographed under UV light.
1.5 Chromosome counting and genomicin situhybridization (GISH)
The root tips were pretreated with 0.002 mol/L 8-hydroxyquinoline at room temperature for 1 h,fixed with 3:1 (V/V) ethyl alcohol: acetic acid, and transferred to absolute alcohol at 4 °C for at least 24 h to confirm the chromosome number of the putative somatic hybrids. They were stored in 70% (V/V)ethanol at 4 °C prior to chromosomal observation.Chromosome preparations were done according to the method described by Lian[14].
Genomic DNA ofB.campestriswas labeled with fluorescein-11-dUTP via the nick translation method using a nick-translation kit (Boehringer-11-Mannheim, Roche, Germany) and was used as a probe. The average fragment length was 100 bp to 200 bp. For the GISH analysis, genomic-blocking DNA was boiled in water containing 10 mol/L NaOH for 40 min to 45 min. The probe and blocking DNA concentrations in the hybridization mixture were 3 and 20 μg/mL, respectively.In situhybridization was performed according to the methods of Leitch[21]. The hybridization signals of theB.campestrisprobes were detected using fluorescein isothiocyanate-anti-avidine (FITC). Chromosomes were counterstained with propidium iodide (Roche,Basel, Switzerland), mounted in an antifade solution(Vector Laboratories, Cambridge, MA, USA), and examined using fluorescence microscopy.
1.6 Morphological comparison
The morphological characteristics, such as leaf shape, size, and flower color, of the protoplast fusion plants were investigated and compared with those of the fusion parents. Putative somatic hybrids were acclimatized on a mixture of peat moss and perlite during flowering induction. When the plant growth reached the 14-leaf stage, plant vernalization was performed at 4 °C for 45 days. After a low-temperature treatment, the plants were placed in a greenhouse under a 24 h illumination (20 Klux).The morphologies and fertilities of the progenies and seed set from the first and second generations were also investigated.
2 Results and discussion
2.1 Protoplast fusion and plant regeneration
In this present experiment, high yield of protoplasts was obtained from healthy germinating seedlings (Fig.1A). The polyethylene glycol(PEG)-mediated protoplasts (Fig.1B) were cultured in 2 mL of the modified K8p medium containing 1 mg/L 2,4-D, 0.5 mg/L 6-BA, 0.1 mg/L NAA, and 1 mg/L Kin. After two days of culture, the first cell division occurred, and the 5- to 7-day cultures were transferred to Kao’s basal medium[22]containing 0.25 mg/L 2,4-D, 0.025 mg/L 6-BA, 0.025 mg/L NAA, and 0.25 mg/L Kin. The medium was supplemented with 6% mannitol and semi-solidified with 0.1% agarose. When microcalli (Fig.1C)reached diameters of 2 mm to 3 mm, they were transferred to a shoot induction medium added with a number of plant growth regulators, namely,0.2 mg/L ZEA, 1 mg/L 6-BA, 0.5 mg/L Kin, and 0.4 mg/L NAA, as well as 3% sucrose and 0.8%agar. After 3 months of proliferation, calli with small greenish nodules produced shoots (Fig.1D). Theproduction of shoots is often associated with root hair formation on the callus. A total of 35 regenerated green plants were obtained from 320 calli, and the plant regeneration frequency was 10.94%. All regenerated plants were transferred to pots, and eleven survived in the greenhouse, they were marked as No.1 to No.11.
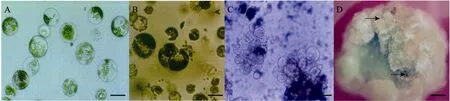
Fig.1 Protoplast isolation, fusion and cell culture. (A) Isolated protoplast from cotyledons (Bar=50 μm). (B) Fused protoplast(Bar=50 μm). (C) Cell division (Bar=50 μm). (D) Regenerated plants from calli (black arrows).Bar=1 mm.
2.1 Ploidy estimation using flow cytometry
Flow cytometry is a useful technique in detecting the variation in the ploidy status among genotypes of the same species, such as that ofB. napus[23], as well as in interspecific hybrids[24-25].Flow cytometry is also widely used to study the genome size and stability of different plant materials culturedin vitro[26-28]. The hybridity of four plants with different chromosome numbers were confirmed using a ploidy analyzer, as previously described by Arumuganathan and Earle[24].
In this study, flow cytometry analysis reveals variations in the ploidy level of the somatic hybrids,and the typical position of the histograms of the fluorescence was obtained.B. campestrisshowed one peak with a diploid at around channel 75(Fig.2A), according to the Partec User Manual. The peak of theB. oleraceadiploid was located at around channel 50 (Fig.2B). The peaks of the four regenerated plants (No.1 to No.4) after the protoplast fusions appeared at the channel near 150,indicating that the tetraploid somatic hybrid was derived from a combination ofB. campestrisandB. oleraceagenomes (Fig.2C). The peaks of another four plants (No.5 to No.8) appeared at the channel 250, indicating that the hexaploid somatic hybrid was regenerated from cell fusion (Fig.2D). The peaks of three putative hybrids (No.9 to No.11)appeared at channel 350, suggesting that the polyploid plant was obtained through protoplast fusion and cell (Fig.2E, F).
2.2 RAPD analysis
The putative somatic hybrids were further confirmed by RAPD analysis using 40 primers on the basis of morphological and chromosomal observations. The profiles of the amplified products from both parents obtained using the OPA-17 (5′-GA CCGCTTGT-3′) primer were clearly polymorphic,and the putative hybrid plants showed specific bands characteristic of the two parents (Fig.3). These results indicate that genetic materials from both parents were successfully incorporated into the somatic hybrids. The RAPD markers were frequently used for characterization of nuclear genomes. Such markers require very small quantities of DNA and generally reveal dominant alleles. The markers allow the polymorphic amplification of each parental genotype, which is necessary to confirm the presence of both genomes in putative hybrids[29].
2.3 Chromosome counting and GISH analysis
All putative somatic hybrids had between 38 and 72 chromosomes. The plants were classified into three types according to the chromosome number, as follows: type I, which included 4 plants (No.1 to No.4), possess 38 chromosomes corresponding to the sum of those ofB. campestrisandB. oleracea(Fig.4A, B); type II, which included 4 plants (No.5 to No.8), have 58 to 60 chromosomes (Fig.4C);and type III, which included 3 plants (No.9 to No.11), have 70 to 76 chromosomes (Fig.4E, F).These results are consistent with those of the flow cytometry analysis.
GISH was conducted on regenerated plants to characterize their chromosome composition. In three regenerants (No.9 to No.11), the signals of variable sizes and intensities were mainly located in the terminal and centromeric parts of some mitotic chromosomes (Fig.4D−F). The DNA of theB.campestris, which exhibits yellow fluorescence,was thoroughly mixed with the hybrid chromosome(Fig.4). GISH inBrassicawas normally characterized by strong signals at centromeric heterochromatin and only very weak hybridization on chromosome arms[30-31], for the low copy numbers of dispersed repeated sequences inBrassicaand related genera.By contrast, in this experiment,B. campestrissignals were distributed in certain parts of the chromosomes (Fig.4A, C, E), or large patches of chromosomes such as in the centromeric (Fig.4B, D, F)and terminal positions (Fig.4B) of chromosomes.No intact chromosomes fromB. campestriswere observedin these regenerants. Similar results were reported in hybrids plants of theBrassicaspecies[32-33].
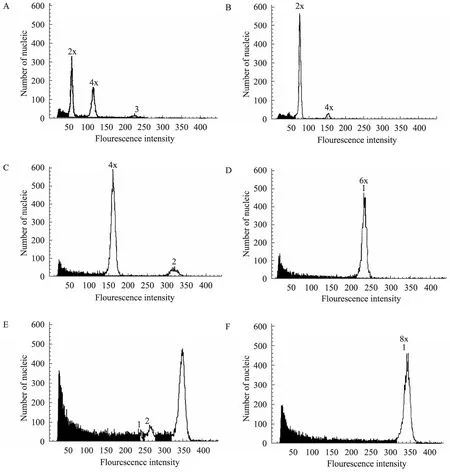
Fig.2 Histogram of the fluorescence intensities for isolated cells from chopping leaves of fusion partners and somatic hybrids. (A)B. campestris(Chinese cabbage). (B)B. oleracea(cabbage).(C) Tetraploid somatic hybrids of No.1. (D) Hexaploid somatic hybrids of No.8. (E, F) Polyploid somatic hybrids of No.9 and No.11.
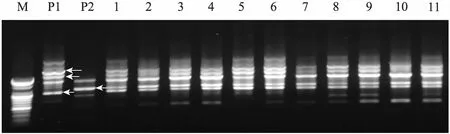
Fig.3 RAPD analysis of parental lines and somatic hybrids. M: DNA marker. P1:B. campestris; P2:B. oleracea; 1–11: somatic hybrids (arrows showB. canpestrisandB. oleraceaspecific bands).
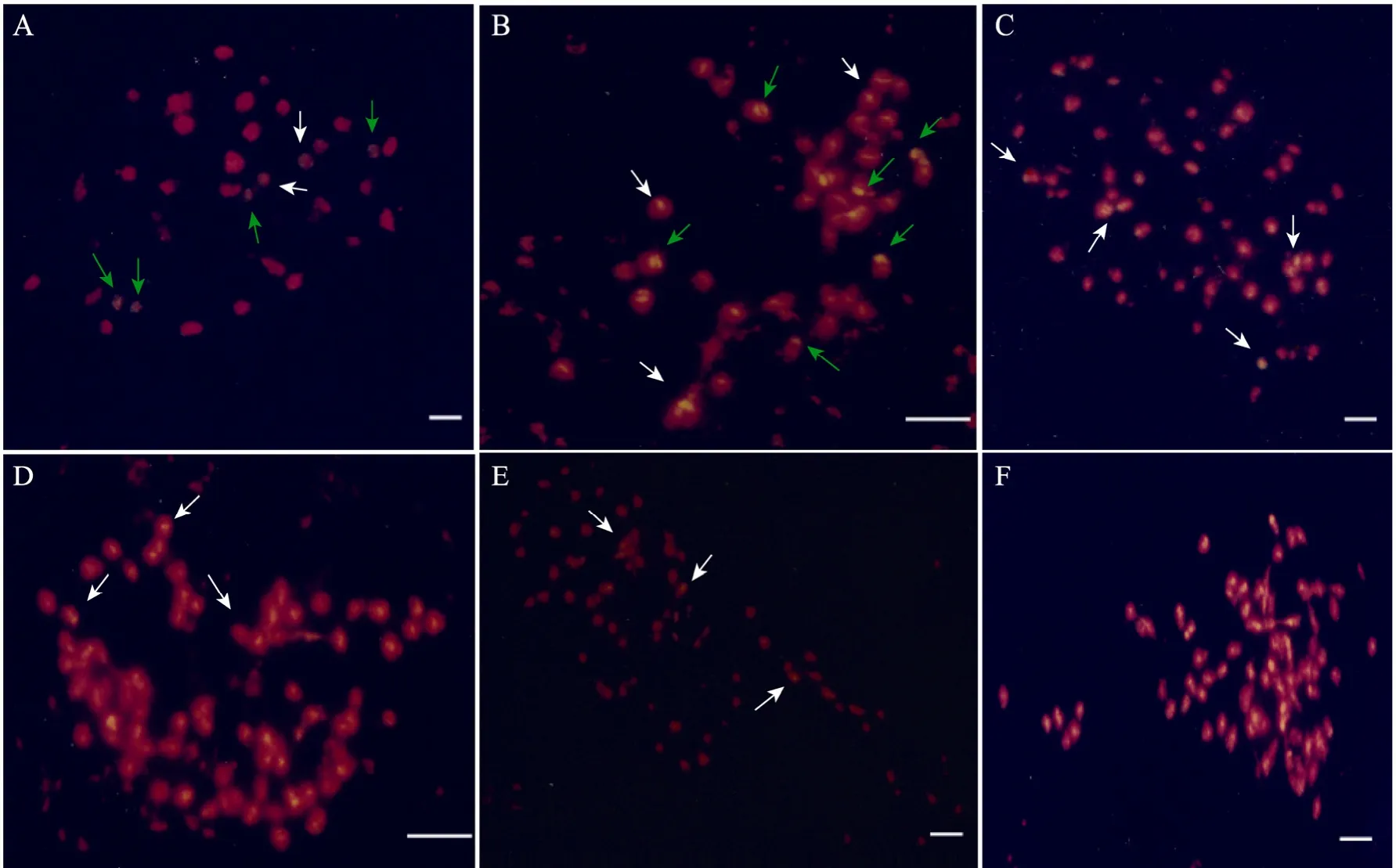
Fig.4 Chromosome counting and genomicin situhybridization of somatic hybrids betweenB.oleraceaandB. campestris. Yellow signals (allows) are from the labelledB.campestrisprobe, chromosome counterstained by propidium iodide (PI). (A, B) Mitotic cells of somatic hybrids of No.1 and No.4 (2n=38),B. campestrissignals were distributed terminal positions (green arrow) or in the centromeric (white arrow) of chromosomes. (C) Mitotic cells of somatic hybrids of No.8 (2n=60),B. campestrissignals were distributed in certain parts (white arrow) of chromosomes. (D) Mitotic cells of somatic hybrids of No.9 (2n=72),B. campestrissignals were distributed large patches of chromosomes. (E) Mitotic cells of somatic hybrids of No.10 (2n=76),B. campestrissignals were distributed in certain parts (white arrow) of chromosomes. (F) Mitotic cells of somatic hybrids of No.11 (2n=76),B. campestrissignals were distributed large patches of chromosomes.Bar=5 μm.
2.4 Morphological characterization of somatic hybrids
The regenerated plants and fusion parents were simultaneously transferred into pots and cultivated in a greenhouse, and their morphological traits werecompared. Wide variations in morphology were observed, especially within the group of plants with abnormal chromosome numbers. The Chinese cabbage has enlarged petioles (Fig.5A-a), whereas cabbage has narrow petioles (Fig.5A-b). The petioles of a somatic hybrid were intermediate to those of the parents (Fig.5A-c). The putative hybrids exhibited varied morphologies compared with those of the regenerated plants and their fusion parents. Four plants with 38 chromosomes showed intermediate leaf morphology (Fig.5A-1 to 4), and seven plants had thicknesses comparable to other regenerants and their fusion parents (Fig.5A-5 to 11).
Four mature plants, whose leaves were crenate/lyrate, deep green, thin, and covered with a waxen powder, were similar to cabbage (Fig.5B-1 to 4). Four plants whose leaves were uncrenated/lyrate, green, with a waxen coating exhibited morphologies intermediate of those of the fusion parents (Fig.5B-5 to 8). Three plants had the most thick, deep green and uncrenated/lyrate leaves with waxen powder, which is similar to cabbage (Fig.5B-11).
After vernalization, all regenerated plants and fusion parents exhibited bolting after two months of cultivation under greenhouse conditions. The floral apex branching patterns were intermediate of those of cabbage and Chinese cabbage with small, loosely branched terminal heads.
2.5 Analysis of the seed set
Some seeds can be obtained from hybrids with more than 38 chromosomes after self-pollination.However, the seed set within the group of hybrids with 38 chromosomes significantly varies. The seed set was measured as the number of seeds/placenta.The somatic hybrids had 0.11 seeds per pod after self-pollination, even though the plants produced numerous pollen grains. The plants produced 0.03 to 0.04 seeds after backcrossing with Chinese cabbage,and had 0.23 to 0.76 seeds per pod after open-pollination (Table 1). Similar results have been reported by Yamagishi[34]and Chen[35]. Thus,somatic hybrids of Chinese cabbage and cabbage have low fertility, as reported by Sundberg[36],probably due to somatic incompatibility. Therefore,the incorporation of the total genomes of two very distantly related species in a hybrid through somatic hybridization has two obvious disadvantages,namely, the introduction of a large amount of exotic genetic material accompanying the expected gene(s),and the genetic imbalance leading to somatic incompatibility[37].

Fig.5 Morphological traits of somatic hybrids. (A) a: the leaf shape of cabbage; b: the leaf shape of Chinese cabbage. No.1 to No.4:the leaf shapes of the regenerants with 38 chromosomes. No.5 to No.8: the leaf shapes of the regenerants with 58 to 60 chromosomes.No.9 to No.11: the leaf shapes of the regenerants with 70 to 76 chromosomes. (B) Regenerated plants from fusion derived calli.Bar=10 cm.
2.6 First generation via self pollination or backcrossing
Reciprocal sexual crosses were performed between two of somatic hybrids with 38 chromosomes and the fusion parents. Moreover, the somatic hybrids were self-pollinated. The progenies obtained from the crosses between the somatic hybrids and one of the fusion parent-Chinese cabbage exhibited Chinese cabbage like morphologies, including trichom density on the leaf surface and yellow flowers; however, purple stems,petiole, and big flowers were also observed. The plants obtained in the first backcrossing generation appeared normal, exhibiting a morphology intermediate of those of the parents. They had branched growth habit, purple or white stems from the somatic hybrids, and leaves with intermediate size and shape (Table 2, 3). All progenies showed bluish waxy leaves, and the leaf color was very similar to that of cabbage. Pollen fertility was gradually recovered in the first and second progenies.
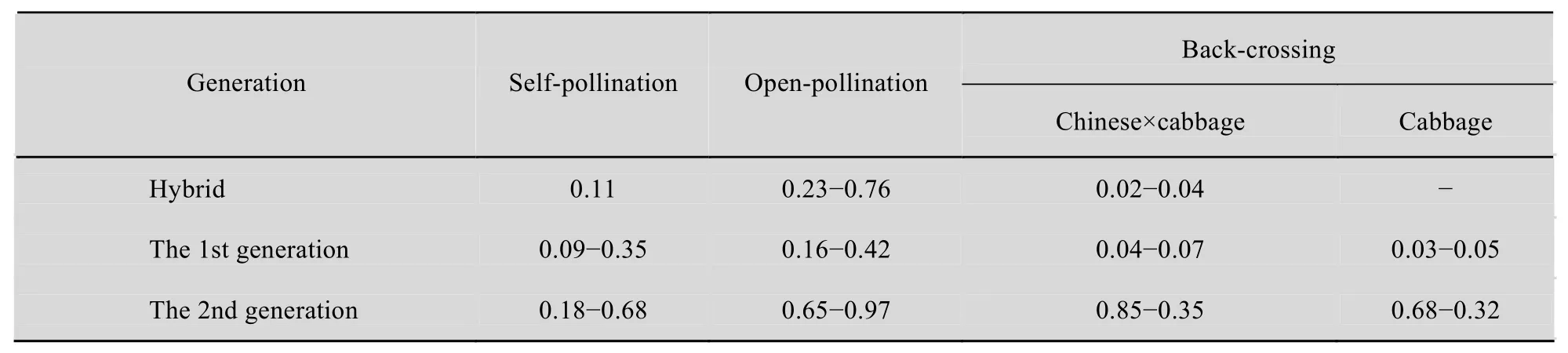
Table 1 The fertility of somatic hybrids and their progenies (Chinese cabbage × cabbage, Unit: seeds/set)
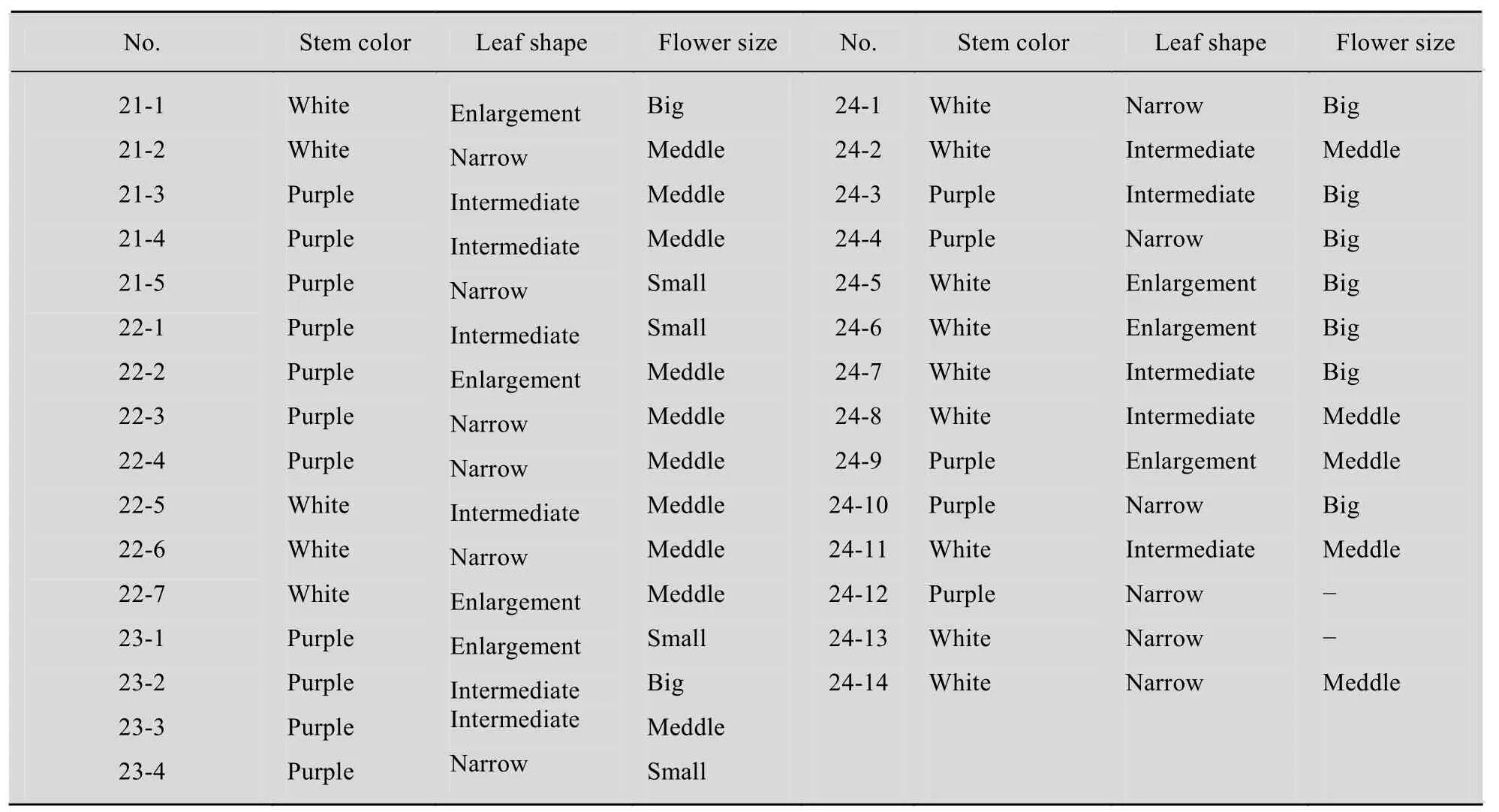
Table 2 Characteristics of the first progenies from one somatic hybrid (SH1)

Table 3 Characteristics of the first progenies from one somatic hybrid (SH2)
2.7 Second backcrossing generation of the somatic hybrids
The BC1 plants were reciprocally crossed withB. campestristo determine the good combinations of crossing parents for the production of BC2 seeds.Based on the morphological characteristics, the progenies obtained from the BC1 seeds showed more suitable variations than those from the first generation. The plants from the first generation,which was similar to cabbage, showed less variation(Fig.6A). However, most of the plants from the second generation, which were similar to Chinese cabbage, showed extensive variation; nevertheless,they still retained some of the Chinese cabbage phenotypes, such as wide petioles (Fig.6B). The progenies of the somatic hybrids from backcrossing had the thick and waxy leaves of cabbage and wide petioles of Chinese cabbage, plants also showed both Chinese cabbage and cabbage morphological features (Fig.6C). Their growth habits were more similar to that of cabbage. The second backcrossing generation showed variable leaf morphology segregation, (Fig.6D–F). With regard to leaf division and marginal incisions of the seedlings, the mature seeds showed undulate/lyrate characteristics.Petioles and midvein enlargement were also intermediate and narrow. Most of the hybrids showed curled-up leaves (Fig.6F).

Fig.6 Plant morphological features of the first and second progenies after crossing with Chinese cabbage. (A) The plants from the first generation. (B,C) The plants derived from the second generation. (D-F) Different shapes of leaves of the second generation plants.Bar = 5 cm.
3 Conclusion
In conclusion, somatic hybrids betweenB. campestrisandB. oleraceaand their backcross progenies were successfully produced, which were characterized by both molecular and cytological analysis. Random amplified polymorphic DNA(RAPD) and Genomicin situhybridization (GISH)techniques were applied to demonstrate the introgressions of two genomes in interspecific somatic hybrids and their progenies.
These interspecific hybrids and their progeny have great potential as a bridge material for the transfer of economically important attributes from cabbage to Chinese cabbage. The agronomical potential of the hybrid progeny is currently under evaluation based on its advancement, improvement,and use.
Acknowledgement The author is grateful to Nam Kwon Baek for providing seed.
REFERENCES
[1] Inomata N. Crossability and cytology of hybrid progenies in the cross betweenBrassica campestrisand three wild relatives ofB. oleracea,B.bourgeaui,B. creticaandB. montana. Euphytica,1993, 69(1/2): 7−17.
[2] Choudhary BR, Joshi P. Crossability ofBrassica tournefortiiandB. rapa, and morphology and cytology of their F1 hybrids. Theor Appl Genet,2001, 102(6/7): 1123−1128.
[3] Waara S, Glimelius K. The potential of somatic hybridization in crop breeding. Euphytica, 1995,85(1/3): 217−233.
[4] Chen FH, Wang H, Li ZY. Production and genetic analysis of partial hybrids in intertribal crosses betweenBrassicaspecies (B. rapa, B. napus) andCapsella bursapastoris.Plant Cell Rep, 2007,26(10): 1791−1800.
[5] Tu YQ, Sun J, Liu Y, Ge XH, et al. Production and characterization of intertribal somatic hybrids ofRaphanus sativusandBrassica rapawith dye and medicinal plant Isatis indigotica Plant Cell Rep,2008, 27(5): 873−883.
[6] Du XZ, Ge XH, Yao XC, et al. Production and cytogenetic characterization of intertribal somatic hybrids betweenBrassica napusandIsatis indigoticaand backcross progenies. Plant Cell Rep,2009, 28(7): 1105−1113.
[7] Dudits D, Maroy E, Praznovszky T, et al. Transfer of resistance traits from carrot into tobacco by asymmetric somatic hybridization: regeneration of fertile plants. Proc Natl Acad Sci USA, 1987,84(23): 8434−8438.
[8] Terada R, Yamashita Y, Nishiyashi S, et al.Somatic hybrids betweenBrassica oleraceaandB.campestris: selection by the use of iodoacetamide inactivation and regeneration ability. Theor Appl Genet, 1987, 73(3): 379−384.
[9] Rosén B, Halldén C, Heneen WK. DiploidBrassica napussomatic hybrids: characterizationof nuclear and organellar DNA. Theor Appl Genet, 1988,76(2): 197−203.
[10] Ozminkowski RH, Jourdan PS. Expression of self-incompatibility and fertility ofBrassica napusL. resynthesized by interspecific somatic hybridization. Euphytica, 1993, 65(2): 153−160.
[11] Cardi T, Earle ED. Production of new CMSBrassica oleraceaby transfer of “Anand”cytoplasm fromB. rapathrough protoplast fusion.Theor Appl Genet, 1997, 94(2): 204−212.
[12] Sigareva MA, Earle ED. Direct transfer of a cold tolerant Ogura male sterile cytoplasm into cabbage(Brassica oleraceassp.capitata) via protoplast fusion. Theor Appl Genet, 1997, 94(2): 213−220.
[13] Lian YJ, Lin GZ, Zhao XM, et al. Production and genetic characterization of somatic hybrids between leaf mustard (Brassica juncea) and broccoli(Brassica oleracea).In VitroCell Dev Biol Plant,2011, 47(2): 289−296.
[14] Lian YJ, Lin GZ, Zhao XM. Morphological,cytological, and molecular characterization ofhybrids and their progenies derived from the somatic hybridization ofBrassica campestrisandBrassica oleracea. Chin J Biotech, 2011, 27(11):1586−1597.
廉玉姬, 林光哲, 赵小梅. 青花菜与白菜间体细胞杂种获得与遗传特性鉴定. 生物工程学报,2011, 27(11): 1586−1597.
[15] Hansen LN, Earle ED. Transfer of resistance ofXanthominascampestrispvcampestrisintoBrassica oleraceaL. by protoplast fusion. Theor Appl Genet, 1995, 91: 1293−1300.
[16] Ren JP, Dickson MH, Earle ED. Improved resistance to bacterial soft rot by protoplast fusion betweenBrassica rapaandB. oleracea. Theor Appl Genet, 2000, 100(15): 810−819.
[17] Murashige T, Skoog F. A revised medium for rapid grown and bio−assays with tobacco tissue cultures.Physiol Plant, 1962, 15: 473−497.
[18] Lian YJ, Lim HT. Plant regeneration ofB. junceathrough plant tissue and protoplast culture. J Plant Biotechnol, 2001, 3: 27−31.
[19] Glimelius K, Mats D, Hugo FF. Selection and enrichment of plant protoplast heterokaryons ofBrassicaceaeby flow sorting. Plant Sci, 1986, 45:133−141.
[20] Doyle JJ, Doyle JI. Isolation of plant DNA from fresh tissue. Focus, 1990, 12: 13−15.
[21] Leitch AR, Schwarzacher T, Jackson D, et al.In situHybridization: A Practical Guide (Microscopy Guide No.27). Oxford: BIOS Scientific Publisher Ltd, 1994.
[22] Kao KN, Michcharyluk MR. Nutritional requirements for growth ofVicia hajastanacells and protoplasts at a very low population density in liquid media. Planta, 1975, 126(2): 105−110.
[23] Takahira J, Cousin A, Nelson MN, et al.Improvement in efficiency of microspore culture to produce doubled haploid canola (Brassica napusL.) by flow cytometry. Plant Cell Tiss Organ Cult,2011, 104(1): 51−59.
[24] Arumuganathan K, Earle ED. Estimation of nuclear DNA content of plants by flow cytometry. Plant Mol Bio Rep, 1991, 9(3): 229−233.
[25] Tiwari JK, Sarkar PD, Pandey SK, et al. Molecular and morphological characterization of somatic hybrids betweenSolanum tuberosumL. andS.etuberosumLindl. Plant Cell Tiss Organ Cult,2010, 103(2): 175−187.
[26] Clarindo WR, de Carvalho CR, Araújo FS et al.Recovering polyploid papayain vitroregenerants as screened by flow cytometry. Cell Tiss Organ Cult, 2008, 92(2): 207−214.
[27] Makowczyńska J, Andrzejewska-Golec E,Sliwinska E. Nuclear DNA content in different plant materials ofPlantago asiaticaL. culturedin vitro. Plant Cell Tiss Organ Cult, 2008, 94(1):65−71.
[28] Mallón R, Rodríguez-Oubiňa J, Gonza´lez ML.In vitropropagation of the endangered plant Centaurea ultreiae: assessment of genetic stability by cytological studies, flow cytometry and RAPD analysis. Plant Cell Tiss Organ Cult, 2010, 101(1):31−39.
[29] Fock I, Collonnier C, Luisetti J, et al. Use of Solanum stenotomum for introduction of resistance to bacterial wilt in somatic hybrids of potato. Plant Physiol Biochem, 2001, 39(10): 899−908.
[30] Snowdon RJ, Köhler W, Friedt W, et al. Genomicin situhybridization inBrassicaamphidiploids and interspecific hybrids. Theor Appl Genet, 1997,95(8): 320−1324.
[31] Wang YP, Snowdon RJ, Rudloff E, et al.Cytogenetic characterization andfae1 gene variation in progenies from asymmetric so somatic hybrids betweenBrassica napusandCrambe abyssinica. Genome, 2004, 47(4): 724−731.
[32] Li ZY, Ceccarelli M, Minelli S, et al. High efficiency production and genomicin situhybridization analysis ofBrassicaaneuploids and homozygous plants. Sci China: Series C, 2003,46(1): 104−112.
[33] Ma N, Li ZY, Cartagena JA, et al. GISH and AFLP analyses of novelBrassica napuslines derived from one hybrid betweenB. napusandOrychophragmus violaceus.Plant Cell Rep, 2006,25(10): 1089−1093.
[34] Yamagishi H, Terachi T. Molecular and biological studies on male-sterile cytoplasm in the Cruciferae.I. The origin and distribution of Ogura male-sterile cytoplasm in Japanese wild radishes (Raphanus sativus L.) revealed by PCR-aided assay of their mitochondrial DNAs. Theor Appl Genet, 1994,87(8): 996−1000.
[35] Chen FH, Wang H, Li ZY. Production and genetic analysis of partial hybrids in intertribal crosses betweenBrassicaspecies (B. rapa, B. napus) andCapsella bursa pastoris.Plant Cell Rep, 2007,26(10): 1791−1800.
[36] Sundberg E. Somatic hybrids and cybrids withinBrassicaceae:studies focused on refining production methods and identifying factors influencing the genetic composition of somatic hybrids[D]. Uppsala: Swedish University of Agricultural Sciences. 1991: 7−44.
[37] Liu JH, Xu XY, Deng XX. Intergeneric somatic hybridization and its application to crop genetic improvement. Plant Cell Tiss Org Cult, 2005,82(1): 19−44.
Production and characterization of a somatic hybrid of Chinese cabbage and cabbage
Yuji Lian
College of Life Science,Linyi University,Linyi276005,Shandong,China
廉玉姬. 白菜与甘蓝之间体细胞杂交种获得与遗传特性鉴定. 生物工程学报, 2012, 28(9): 1080−1092.
Lian YJ. Production and characterization of a somatic hybrid of Chinese cabbage and cabbage. Chin J Biotech, 2012, 28(9):1080−1092.
Received:March 12, 2012;Accepted:July 2, 2012
Supported by:Agricultural R&D Promotion Center, Ministry for Food, Agriculture, Forestry and Fisheries, Republic of Korea (No.204027-02-1-SB010).
Corresponding author:Yuji Lian. Tel: +86-539-8766310; E-mail: lianyuji@lyu.edu.cn.
韩国农林水产部农林技术管理中心项目 (No. 204027-02-1-SB010) 资助。
猜你喜欢
今日农业(2021年14期)2021-11-25
今日农业(2020年24期)2020-12-15
科学(2020年4期)2020-11-26
四川蚕业(2020年2期)2020-07-10
河北果树(2020年1期)2020-02-09
资源再生(2019年1期)2019-03-04
科学导报(2018年47期)2018-05-14
中国环境监察(2016年11期)2016-10-24
中西医结合心脑血管病杂志(2016年20期)2016-03-01
超硬材料工程(2016年1期)2016-02-28
