
hfgl2凝血酶原酶基因5′端调控序列的克隆及生物信息学分析*
2012-09-27 11:20李学军李永敢孙碧红
重庆医学 2012年9期
李学军,张 灏,李永敢,孙碧红
(1.广西壮族自治区人民医院血液科,南宁530021;2.哈尔滨血液病肿瘤研究所血液科 150010)
人纤维蛋白原样蛋白2(human fibrinogen-like protein 2,hfgl2)凝血酶原酶是新近发现的一种促凝因子,它可直接激活凝血酶原启动凝血过程,在功能上相当于因子Ⅹa的作用。研究发现hfgl2凝血酶原酶与机体的一些病理过程有关,如习惯性流产、暴发性肝衰竭及移植排斥反应等[1-4]。最近的研究表明,hfgl2凝血酶原酶的激活参与慢性阻塞性肺病的发生[5]。hfgl2凝血酶原酶可能参与记忆T细胞通过肠道的外渗或迁徙,并介导T细胞对靶细胞的细胞毒作用,通过多种机制进行免疫调节[6-8]。因此,对hfgl2凝血酶原酶基因的深入研究,有助于进一步了解相关疾病的病理生理机制。基因的异常调控常常导致基因在疾病相关细胞中的表达异常,引起相应的病理生理改变,目前对hfgl2凝血酶原酶基因的表达调控机制的研究尚不深入。本研究采用生物信息学软件初步预测了hfgl2凝血酶原酶基因近端启动子序列可能的转录调控位点,并以萤火虫荧光素酶报告基因质粒为载体构建了相应基因序列的报告基因质粒,以进一步研究hfgl2凝血酶原酶基因转录调控机制。
1 材料与方法
1.1 质粒与试剂 萤火虫荧光素酶报告基因载体pGL3-Basic、菌株E.coli DH5α、限制性内切酶KpnⅠ和HindⅢ购自美国Promega公司;LA Taq酶及相关聚合酶链反应(polymerase chain reaction,PCR)试剂、DNA提取试剂盒、DNA连接试剂盒购自日本TaKaRa公司;DNA纯化试剂盒、质粒抽提试剂盒购自美国Omega公司。
1.2 序列分析与结构预测
1.2.1 hfgl2凝血酶原酶基因序列的获得 以human fgl2为关键词,搜索美国国立生物技术信息中心核酸数据库,获得hfgl2凝血酶原酶全长mRNA序列。以hfgl2全长mRNA序列进行基本局部对比排列搜索工具(basic local alignment search tool,BLAST)分析,获取人基因组与其同源的基因序列。取基因组序列中包括第一外显子部分序列在内的1 550bp(转录起始点上游1 464bp至下游86bp)的hfgl2凝血酶原酶基因5′端调控区序列进行序列分析和结构预测。
1.2.2 hfgl2凝血酶原酶基因近端启动子序列的预测 登陆http://rulai.csh1.org/tool/FirstEF/及 http://www.fruitfly.org/seq_tools/promoter.html,利用在线分析软件对hfgl2凝血酶原酶基因5′端调控区序列中可能的启动子进行预测。
1.2.3 胞嘧啶鸟嘌呤二核苷酸(cytosine guanine dinucleotides,CpG)岛预测 登陆http://www.ebi.ac.uk/emboss/cpgplot/,利用在线分析软件 EMBOSS(CpGPlot/CpGReport/Isochore)对hfgl2凝血酶原酶基因5′端调控区序列中可能的CpG岛位置进行预测。
1.2.4 hfgl2凝血酶原酶基因近端启动子序列顺式作用元件的预测 登陆http://www.cbrc.jp/research/db/TFSEARCH.html,利用在线分析软件TFSEARCH对hfgl2凝血酶原酶基因5′端侧翼非翻译区(5′untranslated regions,5-UTR)基因序列的转录因子结合位点进行预测。
1.3 hfgl2凝血酶原酶基因近端启动子序列质粒的构建
1.3.1 引物设计与合成 根据hfgl2凝血酶原酶基因-1 464~+86区域的核苷酸序列,设计扩增该序列的PCR引物。上游引物引入一KpnⅠ酶切位点:5′-GAG GGT ACC CAC GAG GTT CAA ACG TAC TG-3′,下 游 引 物 引 入 一HindⅢ酶切位点:5′-GCA AGC TTC TCT GTT TCA TTG TTT GCC A-3′。引物由TaKaRa公司合成。
1.3.2 质粒的构建 按常规方法分离外周血单个核细胞,应用DNA提取试剂盒参照说明提取细胞基因组DNA,以此DNA为模板,采用LATaq酶扩增相应DNA片段。PCR条件为:95℃预变性5min后加入0.5ULA Taq酶;94℃变性1min,60℃退火1.3min,72℃延伸2min,共30个循环,最后1个循环延伸5min。取5μL PCR产物于1%的琼脂糖凝胶上电泳,观察反应结果。KpnⅠ和HindⅢ双酶切PCR产物及载体pGI3-Basic,回收纯化后,将PCR产物与载体大片段进行连接反应,将得到的质粒命名为pGL3-hfgl2。所构建质粒采用KpnⅠ和HindⅢ双酶切鉴定,将含有酶切鉴定正确的E.coli DH5α送交TaKaRa公司测序。
2 结 果
2.1 hfgl2凝血酶原酶基因近端启动子序列的预测 经过分析,hfgl2凝血酶原酶基因5′端调控区序列中可能存在2处启动子序列,结果见表1。
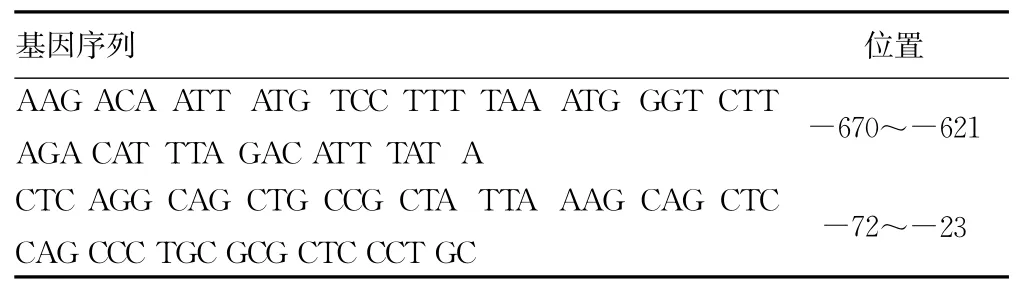
表1 hfgl2凝血酶原酶基因5′端调控区序列启动子序列及位置
2.2 hfgl2凝血酶原酶基因的CpG岛预测结果 在线分析软件EMBOSS(CpGPlot/CpGReport/Isochore)在hfgl2凝血酶原酶基因5′端调控区序列中未发现CpG岛。
2.3 hfgl2凝血酶原酶基因近端启动子序列顺式作用元件的预测 设定阈值大于90分,显示在hfgl2凝血酶原酶基因-1 464~+84区域共有138个、31种转录因子结合位点,其中具有高阈值的转录因子结合位点包括加帽位点(CAP site)、热休克转录因子(heat shock transcription factor,HSF)、尾侧型同源转录因子A(caudal type homeobox transcription factor A,CdxA)、应激反应元件(stress response element,STRE)、Y染色体性别决定区(sex-determining region of Y-chromosome,SRY)、腈 水 解 酶 家 族 成 员 2(nitrilase family,member 2,NIT2)、deltaE、GATA-2、激活蛋白-1(activating protein-1,AP-1)、辅助转录因子p300以及乙醇脱氢酶基因调节因子 (alcohol dehydrogenase gene regulator 1,ADR1)。其中阈值为99分的转录因子结合位点见表2。
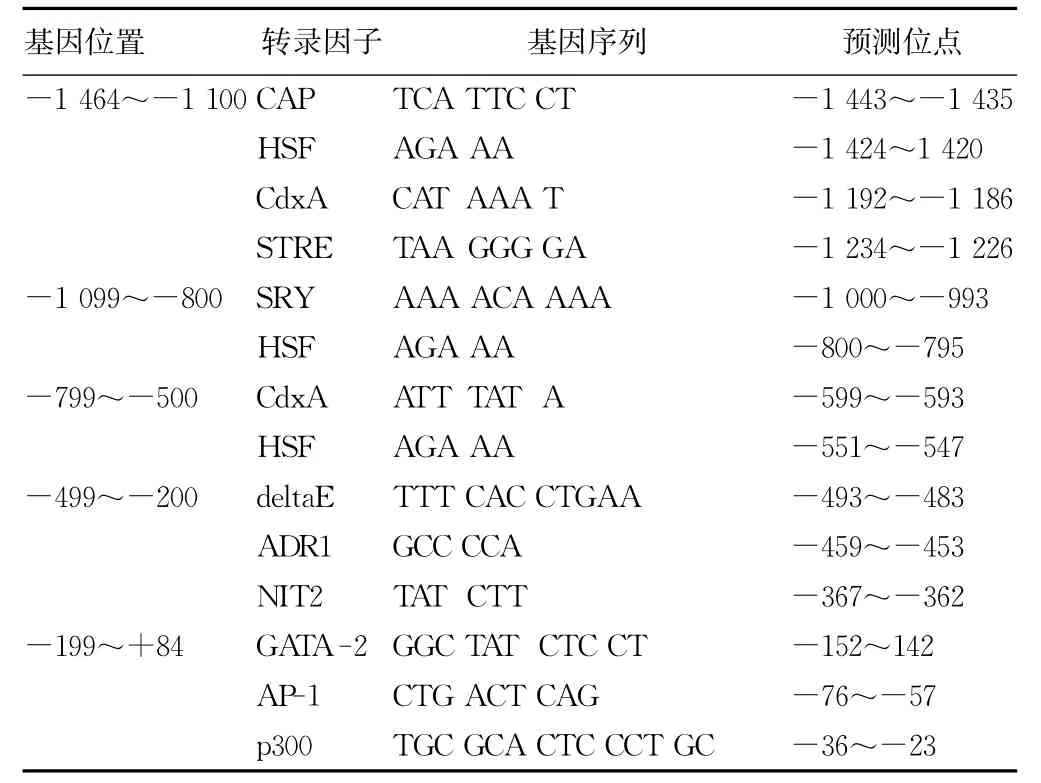
表2 阈值为99分的转录因子结合位点
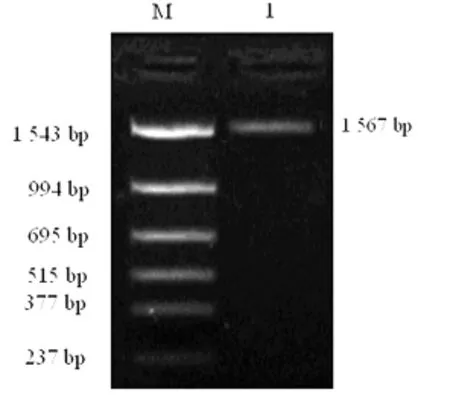
图1 hfgl2凝血酶原酶基因5′端调控区序列PCR扩增电泳图
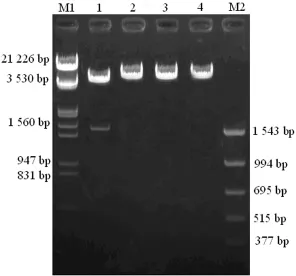
图2 hfgl2凝血酶原酶基因5′端调控区序列质粒酶切图
2.4 重组质粒的克隆与鉴定 以HL-60细胞基因组DNA为模板,PCR扩增获得长度为1 567bp的hfgl2凝血酶原酶基因-1 464~+84序列,与设计长度一致,结果见图1。将PCR扩增基因片段插入pGL3-Basic载体得到重组质粒pGL3-hfgl2,用KpnⅠ和HindⅢ双酶切质粒得到1 560bp大小的DNA片段,与设计的插入序列大小一致,结果见图2。测序结果显示各插入片段与GenBank公布的序列完全一致,见图3。

图3 重组质粒pGL3-hfgl2的部分测序图
3 讨 论
基因表达是指基因在生物体内的转录、剪接、翻译以及转变成具有生物活性的蛋白质分子之前的所有加工过程,转录是基因表达的第一步。转录调控因子,也称为反式作用因子,有序地结合在特定基因5′端侧翼调控序列特殊位点,启动基因的转录和控制基因的转录效率,这些位点即转录因子结合位点或顺式调控元件。每个转录因子的结合位点都有特定的结合模式,找到这些特定的序列片段对研究基因的转录调控有着重要意义[9]。
肿瘤坏死因子α(tumor necrosis factor alpha,TNF-α)、干扰素α(interferonα,IFN-α)等辅助性 T 细胞(helper T cell,Th)1型细胞因子与小鼠及人类自发性流产密切相关,二者单独或联合应用都可诱发小鼠的流产,细胞因子通过触发炎症/血栓形成过程导致流产的发生,推测在人类自发性流产中也存在相同的机制[1,10-12]。在同种异体移植所引起的急性排斥反应中,细胞因子通过激活fgl2凝血酶原酶而介导内皮细胞的损伤,使内皮细胞损伤-凝血-炎症反应-内皮损伤的恶性病理循环发生,导致移植排斥反应[3]。这些细胞因子通过胞膜或细胞内受体经细胞内信号转导系统转换,改变靶基因的转录活性,诱发细胞特定的应答反应。而细胞内受体分布于细胞质或细胞核内,本质上都是配体调控的转录因子,均在细胞核内启动信号转导并影响基因转录。
hfgl2凝血酶原酶基因定位在染色体7q11.23,全长约7kb,有2个外显子,中间有1个长度约2.2kb的内含子。hfgl2凝血酶原酶有2种mRNA转录本,长度分别为4.4kb和1.5kb,其cDNA全长为1 320bp,编码的蛋白质由439个氨基酸组成。通过对鼠血管内皮细胞fgl2凝血酶原酶启动子活性的研究显示,fgl2凝血酶原酶的表达受其近侧启动子序列中的顺式元件调控,在多个反式因子的协同作用下,共同调节内皮细胞的基础表达和对病毒诱导的反应[13-14],其基因5′端调控区存在的正性调控结构域(positive regulatory domain,PRD)与转录密切相关,PRD位于起始位点-87~-39位碱基,研究表明在PRD形成的核蛋白复合物包括特异蛋白(specificity protein,Sp)1/Sp3家族成员、八聚体结合转录因子(octamerbinding transcription factor-1,Oct-1)和 E26转录因子-1(E26 transformation-specific-1,Ets-1)等[15]。本 研 究 显 示,hfgl2 凝血酶原酶基因5′调控区序列中可能存在2处启动子序列,分别位于-670~-621和-72~-23位碱基。在hfgl2凝血酶原酶基因-1 464~+84区域共有138个、31种转录因子结合位点,究竟哪个启动子序列在转录调控中发挥主导作用以及转录因子对启动子有何影响尚有待进一步研究。
CpG岛是基因组中长度为300~3 000bp的富含CpG二核苷酸的一些区域,主要位于基因的启动子和第一外显子区域,常出现在真核生物看家基因(house-keeping gene)的调控区,约有60%以上基因的启动子含有CpG岛。启动子区中CpG岛的未甲基化状态是基因转录所必需的,而CpG序列中胞嘧啶的甲基化可导致基因转录被抑制。本研究通过在线软件分析hfgl2凝血酶原酶基因5′端侧翼调控序列,所分析的区域无CpG岛存在,表明该基因的调控可能不受CpG岛的甲基化影响[5]。
双荧光素酶报告系统是用以研究基因转录活性的,荧光素酶报告基因载体用萤火虫荧光素酶基因作为报告基因,可对启动子及增强子序列作快速及方便的分析,荧光素酶报告基因载体转染到细胞中,可用荧光素酶检测系统灵敏方便地测定荧光素酶基因的表达,pGL3载体含猴空泡病毒40(simian vacuolating virus 40,SV40)启动子及增强子的不同组合,有助于分析DNA片段的转录活性。本实验成功构建了hfgl2凝血酶原酶基因近端启动子转录调控序列荧光素酶报告基因表达载体,为研究该基因5′端侧翼调控序列的各段DNA的调控活性奠定了基础。
[1] Yu G,Sun Y,Foerster K,et al.LPS-induced murine abortions require C5but not C3,and are prevented by upregulating expression of the CD200tolerance signaling molecule[J].Am J Reprod Immunol,2008,60(2):135-140.
[2] Foerster K,Helmy A,Zhu Y,et al.The novel immunoregulatory molecule FGL2:apotential biomarker for severity of chronic hepatitis C virus infection[J].J Hepatol,2010,53(4):608-615.
[3] Ning Q,Sun Y,Han M,et al.Role of fibrinogen-like protein 2prothrombinase/fibroleukin in experimental and human allograft rejection[J].J Immunol,2005,174(11):7403-7411.
[4] Gao S,Wang M,Ye H,et al.Dual interference with novel genes mfgl2and mTNFR1ameliorates murine hepatitis virus type 3-induced fulminant hepatitis in BALB/cJ mice[J].Hum Gene Ther,2010,21(8):969-977.
[5] Liu Y,Xu S,Xiao F,et al.The FGL2/fibroleukin prothrombinase is involved in alveolar macrophage activation in COPD through the MAPK pathway[J].Biochem Biophys Res Commun,2010,396(2):555-561.
[6] Shalev I,Wong KM,Foerster K,et al.The novel CD4+CD25+regulatory T cell effector molecule fibrinogen-like protein 2contributes to the outcome of murine fulminant viral hepatitis[J].Hepatology,2009,49(2):387-397.
[7] Liu H,Shalev I,Manuel J,et al.The FGL2-FcgammaRIIB pathway:a novel mechanism leading to immunosuppression[J].Eur J Immunol,2008,38(11):3114-3126.
[8] Shalev I,Liu H,Koscik C,et al.Targeted deletion of fgl2 leads to impaired regulatory T cell activity and development of autoimmune glomerulonephritis[J].J Immunol,2008,180(1):249-260.
[9] 许丽艳.转录调控的信息学分析[M]//李霞.生物信息学.北京:人民卫生出版社,2010:296-316.
[10]Knackstedt MK,Zenclussen AC,Hertwig K,et al.Th1 cytokines and the prothrombinase fgl2in stress-triggered and inflammatory abortion[J].Am J Reprod Immunol,2003,49(4):210-220.
[11]Hancock WW,Szaba FM,Berggren KN,et al.Intact type 1immunity and immune-associated coagulative responses in mice lacking IFN gamma-inducible fibrinogen-like protein 2[J].Proc Natl Acad Sci U S A,2004,101(9):3005-3010.
[12]Su K,Chen F,Yan WM,et al.Fibrinogen-like protein 2/fib-roleukin prothrombinase contributes to tumor hypercoagulability via IL-2and IFN-gamma[J].World J Gastroenterol,2008,14(39):5980-5989.
[13]Yuwaraj S,Ding J,Liu M,et al.Genomic characterization,localization,and functional expression of FGL2,the human gene encoding fibroleukin:a novel human procoagulant[J].Genomics,2001,71(3):330-338.
[14]Han M,Yan W,Guo W,et al.Hepatitis B virus-induced hFGL2transcription is dependent on c-Ets-2and MAPK signal pathway[J].J Biol Chem,2008,283 (47):32715-32729.
[15]Ning Q,Lakatoo S,Liu M,et al.Induction of prothrombinase fgl2by the nucleocapsid protein of virulent mouse hepatitis virus is dependent on host hepatic nuclear factor-4alpha[J].J Biol Chem,2003,278(18):15541-15549.
猜你喜欢
中国药学药品知识仓库(2022年2期)2022-03-23
天津医科大学学报(2021年4期)2021-08-21
中日友好医院学报(2021年1期)2021-04-14
昆明医科大学学报(2020年12期)2021-01-26
山东医药(2020年9期)2020-05-20
中国药理学通报(2015年2期)2016-01-12
热带农业科学(2015年9期)2015-10-14
中国医学科学院学报(2015年5期)2015-03-01
中国当代医药(2015年30期)2015-03-01
中国当代医药(2015年9期)2015-03-01
