
富马酸亚铁对虹鳟幼鱼生长性能、血液生化指标及微量元素含量的影响
2012-09-20 00:32冯德智张利民王际英李宝山马晶晶蒋锦坤张德瑞姜柯君
动物营养学报 2012年4期
冯德智 张利民 王际英 李宝山 马晶晶 蒋锦坤 张德瑞 姜柯君
(1.上海海洋大学水产与生命学院,上海 201306;2.山东省海洋水产研究所,烟台 264006)
铁是维持机体正常生理功能,保证血红蛋白(Hb)、肌红蛋白、细胞色素和很多酶系统正常功能的必需微量元素。铁还参与氧的运输、DNA合成及电子传递等一系列生化过程[1]。饲料中添加一定量的铁可以提高棉籽粕替代鱼粉的比例,降低肝脏中棉酚的含量[2-3]。鱼类从水和饵料中都可获得所需的铁,但自然水体中可溶性铁浓度很低,无法满足鱼体正常需求,所以饵料是鱼类铁的主要来源[4-5]。研究证明,饲料中缺铁可导致鱼类生长减缓,饲料利用率和存活率降低,组织铁含量下降[6-10]。饲料中铁含量过高会造成脂肪的氧化和酸败,二十碳五烯酸(EPA)、二十二碳六烯酸(DHA)及生育酚损失,影响饲料的稳定性[11-12];此外,铁含量过高会影响抗坏血酸的消化吸收[4],也可产生超氧化物阴离子进而产生有毒性的羟基[13]且能引起鱼体的溃烂[14]。有机形式的矿物元素比无机形式的矿物元素有更高的生物利用率[15]。大西洋鲑(Salmo salar L.)上的研究表明,血红素铁利用率显著高于单质铁和硫酸亚铁[4]。在鲤鱼(Cyprinus carpio)的试验中发现,富马酸亚铁比硫酸亚铁更能提高鲤鱼生长性能和抗病力[16]。不同价态的铁在同一种动物体内表现出不同的生物利用率[10]。肠道黏膜是铁吸收的主要部位[17],且主要以二价铁的形式吸收[18]。罗非鱼(Oreochromis niloticus×O.aureus)的研究中发现,柠檬酸铁是硫酸亚铁效率的1/2[10],因此,本试验选择有机形式的二价铁富马酸亚铁作为添加铁源。饲料中添加铁对虹鳟(Oncorhynchus mykiss)的影响已有相关报道[11,18],但侧重于铁作为强还原剂对饲料稳定性的影响和铁对虹鳟生理调节的影响,虹鳟铁需要量研究尚未见报道。虹鳟饲料中铁的添加量通常是根据经验来添加[11],并无科学指导,仍需试验确定虹鳟准确的铁需要量[18]。本试验通过在饲料中添加不同含量富马酸亚铁,研究铁对虹鳟生长、血液生化指标和微量元素的影响,进而确定虹鳟幼鱼铁的需要量,以期为虹鳟配合饲料的制作和铁代谢的生理生化研究提供基础数据。
1 材料与方法
1.1 试验设计及饲料
选择体质健壮、体重(89.2±0.2)g的虹鳟幼鱼360尾,随机分成6组,每组3个重复,每个重复20尾鱼,以酪蛋白和明胶为蛋白质源,鱼油为脂肪源,配制粗蛋白质含量为42.48%、粗脂肪含量为18.50%的基础饲料。以富马酸亚铁(C4H2FeO4)作为铁源,实测纯度99%,分别在每千克基础饲料中添 加 0、60.71、121.43、242.86、485.71 和1 457.14 mg富马酸亚铁(含铁 0、20、40、80、160和480 mg),以富马酸亚铁替代相同质量的羧甲基纤维素钠,配制成6种精制试验饲料,依次为D0、D20、D40、D80、D160 和 D480组,D0 组为对照组。试验饲料组成及营养水平见表1。经原子吸收分光光度计(AA800型,PerkinElmerTM)测得饲料中实际铁含量分别为 62.60、79.50、99.60、139.30、215.20和538.40 mg/kg。配制饲料时,所有原料粉碎过80目筛,逐级扩大混匀后经螺旋挤压机加工成直径4 mm的硬颗粒饲料,经70℃烘干后放于阴凉干燥处保存、备用。
1.2 试验动物及饲养管理
饲养试验在山东省海洋水产研究所淡水循环养殖系统中进行。试验开始前,虹鳟幼鱼暂养30 d,摄食基础饲料,待其完全适应精制饲料和试验条件后,选择体质健壮、体重(89.2±0.2)g的虹鳟幼鱼,分别放养于18个圆柱形玻璃钢养殖桶中(直径60 cm,高80 cm,水深50 cm)。养殖试验持续60 d,每天投喂2次(08:00,16:30),日投喂量为体重的1.5% ~2.0%,投喂结束30 min后从系统自带的排水口将残饵排出,计算残饵量。试验期间控制水温(14.5±0.5)℃,溶氧>8 mg/L,氨氮<0.4 mg/L。
1.3 样品收集
生长试验结束后,饥饿24 h,称每桶鱼总重,计算增重率。用间氨基苯甲酸乙酯甲磺酸盐(MS-222)麻醉后,随机从每桶中取14尾鱼,称体重,量体长,其中2尾作全鱼用于微量元素测定,剩余12尾尾静脉采血,用于收集血清和制备全血。取血后的鱼分离内脏、肝胰脏、消化道后称重,计算肝体比和脏体比,并取背肌用于微量元素测定。整个操作在冰盘中进行,采样完毕,将样品放入-70℃的超低温冰箱中保存,待测。解剖后的鱼体放入微波炉中加热分离脊椎骨,用超纯水冲洗后,105℃烘干2 h,烘干后的脊椎骨样品粉碎后以乙醚抽提12 h去除脂肪并再次烘干,用于微量元素含量的测定。
1.4 测定指标
增重率(WGR,%)=[(Wt-W0)/W0]×100;
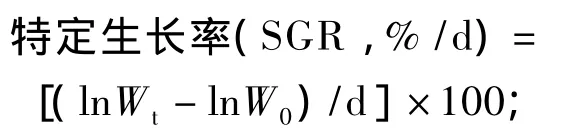
饲料系数(FCR)=F/(Wt-W0);
肝体比(HSI,%)=(肝脏质量/Wt)×100;
脏体比(VSI,%)=(内脏质量/W)t×100;
肥满度(CF,%)=(体重/体长3)×100;
存活率(SR,%)=(成活尾数/总尾数)×100。
式中:W0为试验初鱼体重量(g),Wt为试验末鱼体重量(g),F为摄食干饲料重(g),d为养殖天数。
红细胞数量(RBC)采用血球计数板计数,血细胞比容(Hct)用微量血细胞比容法,血红蛋白、溶菌酶(LZM)和过氧化氢酶(CAT)均采用南京建成生物工程研究所的试剂盒进行测定,铁、锌和铜含量先用微波消解仪(ETHOST)消解,再用原子吸收光谱仪(AA800型,PerkinElmerTM)测定。
1.5 数据统计
采用SPSS 13.0软件进行单因素方差分析(One-way ANOVA),当处理之间差异显著(P<0.05)时,用Duncan氏检验进行多重比较分析,数据用平均值±标准差表示。血红蛋白及肝脏铁含量与饲料铁水平之间的相关关系均采用折线回归分析法。

表1 试验饲料组成及营养水平(干物质基础)Table1 Composition and nutrient levels of experimental diets(DM basis) %
2 结果
2.1 饲料铁水平对虹鳟生长性能的影响
由表2可见,饲料中不同的铁水平对虹鳟幼鱼的增重率、特定生长率、饲料系数、存活率、肝体比、脏体比和肥满度均无显著影响(P>0.05)。
2.2 饲料铁水平对虹鳟血液指标的影响
由表3可见,随着饲料中铁含量的增加,血红蛋白和红细胞数量呈现先上升后平稳的趋势,D40、D80、D160和 D480组显著高于 D0和 D20组(P<0.05),而4组之间差异不显著(P>0.05)。各组间血细胞比容没有显著差异(P>0.05)。以血红蛋白为参考指标,通过折线模型回归分析,虹鳟幼鱼铁需要量为99.8 mg/kg饲料(图1)。
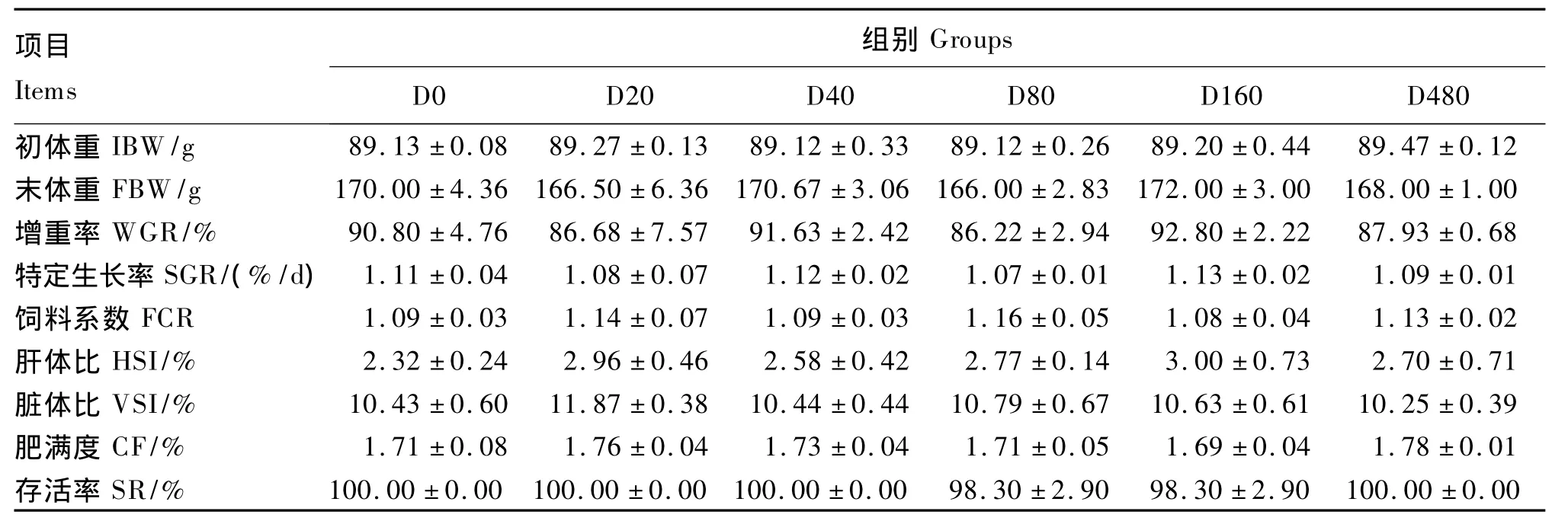
表2 饲料铁水平对虹鳟生长性能和饲料利用的影响Table2 Effects of dietary iron level on growth performance and feed utilization of rainbow trout(n=3)
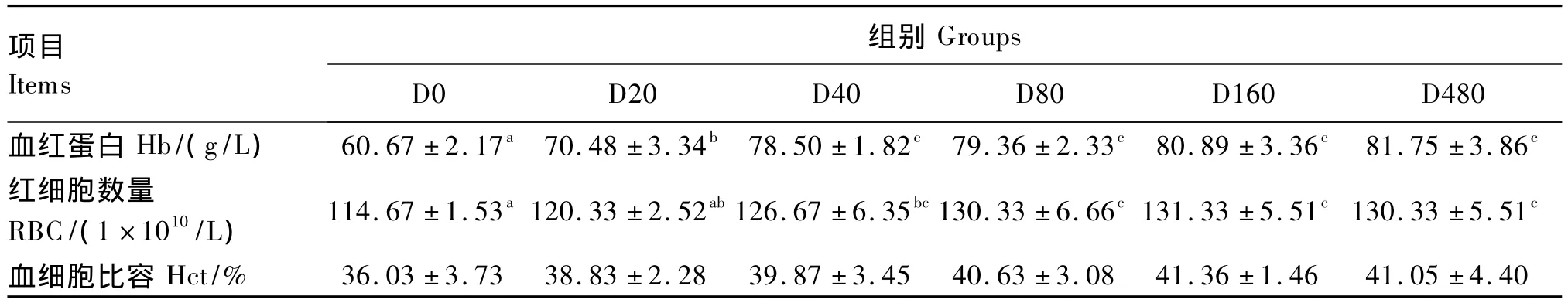
表3 饲料铁水平对虹鳟血液指标的影响Table3 Effects of dietary iron level on blood parameters of rainbow trout(n=3)

图1 饲料铁水平对虹鳟血红蛋白含量的影响Fig.1 Effects of dietary iron level on hemoglobin content of rainbow trout
2.3 饲料铁水平对虹鳟全鱼及组织中铁、锌和铜含量的影响
由表4可见,全鱼铁含量随着饲料中铁水平的升高呈上升趋势。脊椎骨和肌肉铁含量随着饲料中铁水平的升高而逐渐升高,D0和D20组铁含量显著低于其他各组(P<0.05),而两者之间无显著差异(P>0.05)。肝脏铁含量随着饲料中铁水平的增加呈现先上升后平稳的趋势,D40、D80、D160和D480组显著高于D0和D20组(P<0.05),而4组之间差异不显著(P>0.05)。以肝脏铁为参考指标,采用折线回归分析可得,虹鳟幼鱼铁需要量为100.4 mg/kg饲料(图2)。
饲料铁水平对全鱼锌含量影响不显著(P>0.05),但D0组脊椎骨和肌肉锌含量显著高于其他各组(P<0.05)。D0和D20组肝脏锌含量显著高于 D40、D80、D160和 D480组(P <0.05),而两者之间差异不显著(P>0.05)。
全鱼铜含量随着饲料中铁水平的升高呈现逐渐上升的趋势。饲料铁水平对脊椎骨铜含量影响不显著(P>0.05)。肌肉铜含量随着铁水平的升高出现先上升后平稳的趋势,在D40组达到平台,D0组显著低于其他各组(P<0.05)。
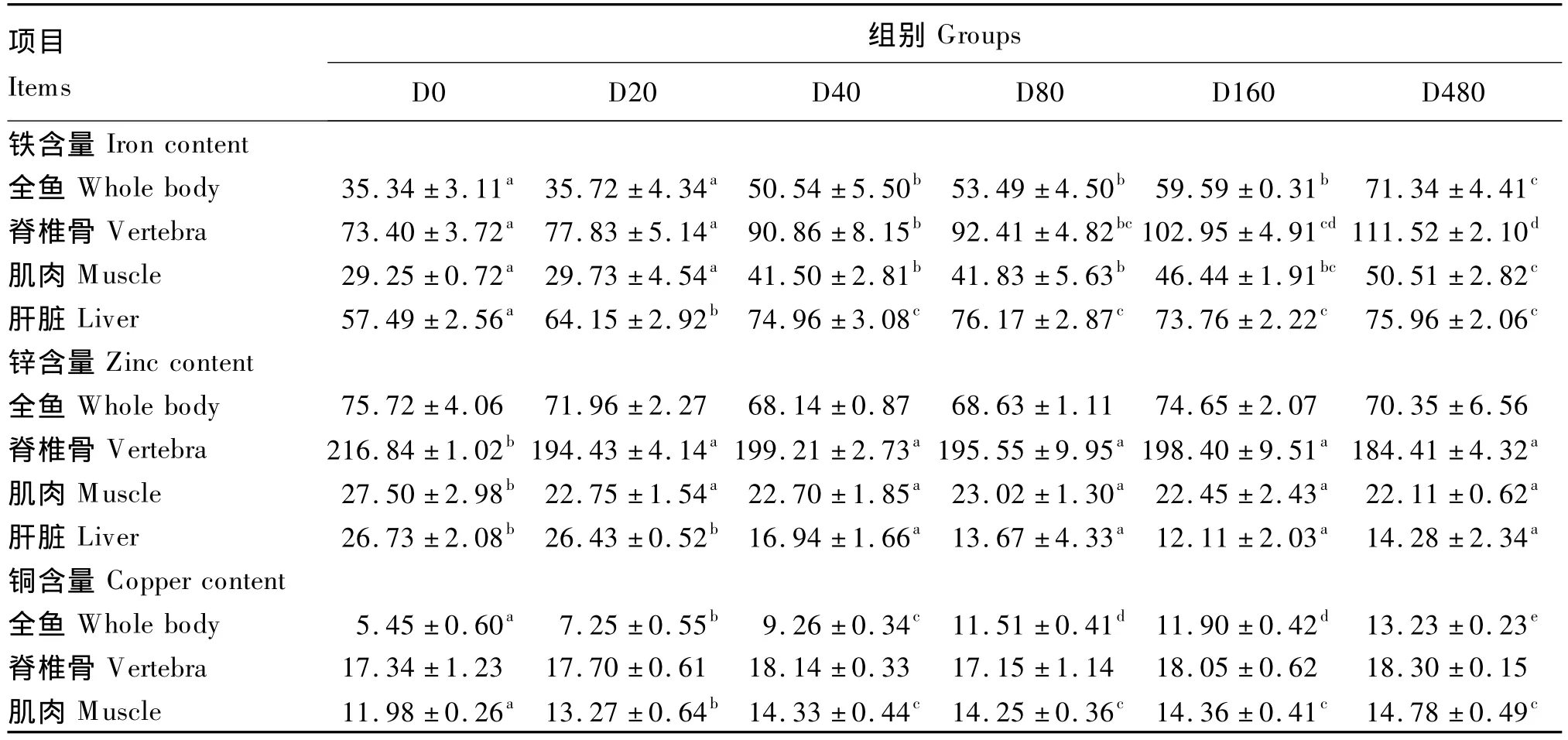
表4 饲料铁水平对虹鳟全鱼和组织铁、锌和铜含量的影响Table4 Effects of dietary iron level on contents of iron,zinc and copper of whole body and tissues of rainbow trout(n=3) mg/kg
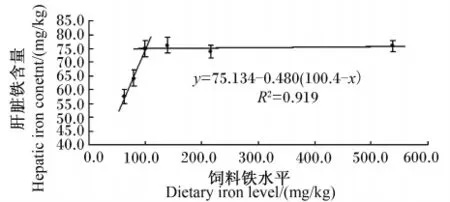
图2 饲料铁水平对虹鳟肝脏铁含量的影响Fig.2 Effects of dietary iron level on hepatic iron content of rainbow trout
2.4 饲料铁水平对血清溶菌酶和过氧化氢酶活性的影响
由表5可见,D0组溶菌酶活性显著低于D20、D40、D80和 D160组(P<0.05),但与 D480组差异不显著(P>0.05)。过氧化氢酶活性随着铁添加量的增加出现先上升后下降的趋势,在D40组达到最大(P<0.05)。
3 讨论
3.1 饲料铁水平对虹鳟生长性能的影响
本试验研究表明,饲料中不同铁添加量对虹鳟幼鱼的增重率、特定生长率、饲料效率、和存活率等无显著影响,这与金头鲷(Sparusaurata)[19]、大西洋鲑[4,20]、溪红点鲑(Salvelinus fontinalis)[21]、鲤鱼[22]、真鲷(Chrysophrys major)[23]和南美白对虾(Penaeus vannamei)[24]等的研究结果相一致,Rigos等[19]认为不能用生长性能来作为评价金头鲷饲料铁添加量的标准,Andersen等[20]指出生长性能并不完全适宜用于评价矿物元素的需要量。然而,对 石 斑 鱼(Epinephelus coioides)[7]、罗 非鱼[10]、中华鳖(Pelodiscus sinensis)[25-26]、斑点叉尾(Ictalurus punctatus)[6,27]、建 鲤 (Cyprinus carpio var.Jian)[28]、鲈 鱼 (Lateolabrax japonicus)[29]、大黄鱼(Pseudosciaena crocea R.)[29]和军曹鱼(Rachycentron canadum)[30]的研究均显示未添加铁饲料组出现鱼类生长抑制现象,其中增重率或特定生长率在罗非鱼[10]、中华鳖[26]、鲈鱼[29]和大黄鱼[29]中用于评价铁需要量。产生以上不同结果的原因可能是基础饲料组铁含量已经满足鱼类生长的需求,也可能是铁对鱼类生长具有一定的种属特异性。试验鱼的大小、投喂次数、投喂环境等诸多因素也会造成上述不同结果。有研究指出,斑点叉尾鮰存活率在饲喂缺铁饲料达13 周以上时明显下降[8,31],而本试验期为 60 d,虹鳟生长无显著性差异可能与试验周期较短有关,具体关系还有待进一步确认。

表5 饲料铁水平对虹鳟血清溶菌酶和过氧化氢酶活性的影响Table5 Effects of dietary iron level on activities of serum lysozyme and catalase of rainbow trout
3.2 饲料铁水平对虹鳟血液学指标的影响
血液指标是通常被用来检测鱼类是否缺铁的重要指标。饲料中铁水平过低会造成贫血,检测贫血的重要指标是血红蛋白[4]。本试验血红蛋白和红细胞数量随着饲料铁水平的升高呈现先上升后平稳的趋势,说明基础饲料中铁含量偏低,难以维持血液指标的正常,还需要向基础饲料中额外添加铁,这与斑点叉尾[6]、真鲷[9,23]、罗非鱼[10]、大西洋鲑[20]和中华鳖[25-26]的研究结果类似。以血红蛋白作为评价指标,通过折线模型分析得出虹鳟幼鱼铁需要量为99.8 mg/kg,这与大西洋鲑(60 ~100 mg/kg)[20]、鲈鱼(95.2 mg/kg)[29]和大黄鱼(101.2 mg/kg)[29]铁需要量相吻合,高于斑点叉尾(30 mg/kg)[6]的铁需要量,但低于真鲷(150 mg/kg)[23]、中华鳖(266 ~ 325 mg/kg)[25]、罗非鱼(150 mg/kg)[10]和建鲤(147.4 mg/kg)[28]的铁需要量,这可能与鱼体种类和大小等因素有关。饲料中添加不同价态或剂型的铁源也是铁需要量出现差异的重要原因,大西洋鲑对单质铁需要量高于硫酸亚铁,对硫酸亚铁需要量高于血红素铁[4],真鲷对柠檬酸铁的需要量高于氯化亚铁和氯化铁[9],罗非鱼对柠檬酸铁的需要量高于硫酸亚铁[10]。
3.3 饲料铁水平对虹鳟全鱼及组织中铁、锌和铜含量的影响
全鱼铁含量随着饲料中铁水平的升高呈上升趋势。D0和D20组全鱼铁含量显著低于D40、D80和D160组,说明D0和D20组饲料铁水平可能已经不足以应对机体对铁的需求,需要动用身体原先储备的铁来满足机体正常的生理功能[29],因此造成铁含量的降低。饲料铁水平偏低的D0和D20组全鱼铁含量明显降低,与大西洋鲑[4]和石斑鱼[7]的研究结果相一致。D480组全鱼铁含量继续升高,显著高于其他各饲料组,可能是吸收的铁在各组织间转移的结果。Carriquiriborde等[18]在研究铁代谢时发现,提高饲料铁水平会促进组织中铁的积累,尤其引起胃、肠道和血液中铁水平的提高。虽然肝脏作为铁储存器官,但过多的铁会转移到胃和肠道中,造成高铁饲料组肠道铁含量是正常组的13倍之多,全鱼铁含量是对照组铁含量的3.7倍。
肝脏是鱼类储存铁的主要器官,随着饲料中铁添加量的增加而逐渐升高[32-33]。肝脏中铁含量已经广泛用于评价铁需要量,在石斑鱼[7]、罗非鱼[10]和中华鳖[25-26]中均有相关报道。本试验肝脏铁含量呈现先上升后平稳的趋势,符合折线模型应用条件,因此用折线模型回归分析得出虹鳟幼鱼铁需要量为100.4 mg/kg,与用血红蛋白作为评价指标时得出的结果相近,与石斑鱼(100 mg/kg)[7]的研究结果也类似。
饲料铁水平的改变对其他金属元素的代谢有明显的影响[34]。铁与锌都是二价离子,在肠道内竞争性吸收,并占用部分共同的转运通道[35]。食用缺锌或高铁的食物都会出现可循环锌水平降低[36]。Carriquiriborde 等[18]指出低铁饲料组肌肉锌水平在第4周出现上升。Ogino等[37]研究虹鳟锌需要量时发现,不额外添加锌饲料组全鱼、脊椎骨、肠道和肝脏铁水平明显高于添加锌饲料组。本试验中D0组脊椎骨和肌肉锌含量明显高于其他各组,D0和D20组肝脏锌含量也显著高于其他各组,说明铁与锌是拮抗关系,与前人研究结果相一致。
铜作为造血反应的促媒,催化铁合成血红蛋白和帮助生成红细胞。铜也参与铁的转运,参与生成铜蓝蛋白(又称亚铁氧化酶),可以将二价铁氧化为三价铁结合到去铁转铁蛋白上合成转铁蛋白[38]。Lanno 等[39]指出随着饲料中铜添加量的增加,肝脏中铜和铁水平都相应的增加。本试验中发现随着饲料中铁水平的增加,全鱼和肌肉中铜含量也显著增加,可能是铁水平增加需要参与转运铁的铜(如铜蓝蛋白)也相应增加的缘故。然而,Ye等[7]发现饲料铁水平对石斑鱼肝脏中铜含量并没有显著影响,Conrad等[40]报道二价铁和三价铁都不共用铜的运输通道,但是Gunshin等[36]指出DCT1(一种金属离子转运体)可以共同转运铁和铜。综上所述,铁与铜的关系还有待进一步研究。
3.4 饲料铁水平对虹鳟血清溶菌酶和过氧化氢酶活性的影响
溶菌酶是一项非特异性免疫指标,可以降低微生物病菌的活力,裂解细胞壁上的肽聚糖,造成病菌微生物的溶解,提高鱼体的抗感染能力。研究指出,一定程度上血清中溶菌酶活性变化与白细胞数目变化相一致,白细胞数目多溶菌酶活性就强,二者呈正相关[41]。Hoepelman 等[42]指出铁可以修复白细胞多形核粒细胞,所以适当的铁水平会提高溶菌酶活性。本试验中D0组溶菌酶活力显著低于D20、D40、D80和D160组,说明随着饲料铁水平的增加溶菌酶活性提升,这与美国红鱼(Sciaenops ocellatus)[43]的研究结果一致。在斑点叉尾鮰饲料中额外添加180 mg/kg铁,斑点叉尾死亡率提高[44]。本试验D480组溶菌酶活性出现略微的下降,但与添加铁试验组差异不显著,可能铁水平过高会提高微生物病菌感染的风险[45]。
过氧化氢酶能催化过氧化氢产生氧气(O2)和水(H2O),是细胞抗氧化防御体系的重要组成部分[33]。铁是保持过氧化氢酶活性必不可少的辅助因子[29]。随着饲料铁水平的提高,过氧化氢酶呈现先上升后下降的趋势,与牙鲆(Paralichthys olivaceus)[46]、大黄鱼和鲈鱼[29]的研究结果一致。有研究表明,缺铁性贫血组氧化物活性显著高于正常组,抗氧化活性和过氧化氢酶活性显著低于正常组,在补充铁后各项指标恢复正常[47],本试验中D0、D20和D40组过氧化氢酶活性随着饲料铁水平的升高逐渐升高,与前人结果一致。抗坏血酸在铁代谢中作为还原剂保证二价铁的高效吸收和利用[31]。有研究指出,饲料铁水平的提高会导致体内维生素C含量明显下降[4],同时还有研究表明维生素C含量低的饲料组血清过氧化氢酶活性明显降低[48],维生素C利用率的不同也会造成过氧化氢酶活性的差异[33],所以本试验中过氧化氢酶在D40~D480组出现下降现象可能是饲料中铁水平的升高致使体内维生素C含量下降或利用率降低造成过氧化氢酶活性下降,具体机理还有待进一步研究。
4 结论
①饲料中添加富马酸亚铁对虹鳟幼鱼生长性能无显著影响,但适量添加可以提高血红蛋白、血细胞比容和红细胞数量,保证全鱼和组织中铁、锌和铜含量处于相对稳定的状态,提高血清溶菌酶和过氧化氢酶活性,从而增强机体免疫力。
②本试验条件下,以血红蛋白和肝脏铁含量为评价指标,虹鳟幼鱼对饲料中铁需要量分别为99.8和100.4 mg/kg。
[1]ANDREW T M,DALNA B,GLADYS O,et al.An iron-regulated ferric reductase associated with the absorption of dietary iron[J].Science,2001,291(2):1755-1759.
[2]LIM S J,LEE K J.Partial replacement of fish meal by cottonseed meal and soybean meal with iron and phytase supplementation for parrot fish Oplegnathus fasciatus[J].Aquaculture,2009,290:283-289.
[3]LIM S J,LEE K J.Supplemental iron and phosphorus increase dietary inclusion of cottonseed and soybean meal in olive flounder(Paralichthys olivaceus)[J].Aquaculture Nutrition,2008,14(5):423-430.
[4]ANDERSEN F,LORENTZEN M,WAAGBØ R,et al.Bioavailability and interactions with other micronutrients of three dietary iron sources in Atlantic salmon,Salmo salar,smolts[J].Aquaculture Nutrition,1997(3):239-246.
[5]National Research Council.Nutrient requirement of fish[S].Washington,D.C.:National Academy Press,1993.
[6]GATLIN D M,WILSON R P.Characterization of iron deficiency and the dietary iron requirement of fingerling channel catfish[J].Aquaculture,1986,52:191-198.
[7]YE C X,LIU Y J,MAI K S,et al.Effect of dietary iron supplement on growth,haematology and microelements of juvenile grouper,Epinephelus coioides[J].Aquaculture Nutrition,2007,13:471-477.
[8]LIM C,KLESIUS P H.Responses of channel catfish(Ictalurus punctatus)fed iron-deficient and replete diets to Edwardsiella ictaluri challenge[J].Aquaculture,1997,157:83-93.
[9]SAKAMOTO S,YONE Y.Availabilities of three ironcompounds as dietary iron sources for red sea bream[J].Bulletin of the Japanese Society of Scientific Fisheries,1979,45(2):231-235.
[10]SHIAU S Y,SU L W.Ferric citrate is half as effective as ferrous sulfate in meeting the iron requirement of juvenile tilapia,Oreochromis niloticus× O.aureus[J].The Journal of Nutrition,2003,133:483-488.
[11]DESJARDINS L M,HICKS B D,HILTON J W.Iron catalyzed oxidation of trout diets and its effect on the growth and physiological response of rainbow trout[J].Fish Physiology and Biochemistry,1987,3(4):173-182.
[12]SUTTON J,BALFRY S,HIGGS D,et al.Impact of iron-catalyzed dietary lipid peroxidation on growth performance,general health and flesh proximate and fatty acid composition of Atlantic salmon(Salmo salar L.)reared in seawater[J].Aquaculture,2006,257:534-557.
[13]SILVA D M,ASKWITH C C,KAPLAN J.Molecular mechanisms of iron uptake in eukaryotes[J].Physiological Reviews,1996,76:31-47.
[14]SALTE R,RØRVIK K A,REED E,et al.Winter ulcers of the skin in Atlantic salmon,Salmo salar L.pathogenesis and possible aetiology[J].Journal of Fish Diseases,1994,17:661-665.
[15]RIDER S A,DAVIES S J,JHA A N,et al.Bioavailability of co-supplemented organic and inorganic zinc and selenium sources in a white fishmeal-based rainbow trout(Oncorhynchus mykiss)diet[J].Journal of Animal Physiology and Animal Nutrition,2010,94:99-110.
[16]张杰,张锦秀,李书伟,等.不同铁源对鲤鱼生长性能和死亡率的影响[J].中国畜牧兽医,2010,37(8):18-20.
[17]TAKESHI W,VISWANATH K,SHUICHI S.Trace minerals in fish nutrition[J].Aquaculture,1997,151:185-207.
[18]CARRIQUIRIBORDE P,HANDY R D,DAVIES S J.Physiological modulation of iron metabolism in rainbow trout(Oncorhynchus mykiss)fed low and high iron diets[J].Journal of Experimental Biology,2004,207:75-86.
[19]RIGOS G,SAMARTZIS A,HENRY M,et al.Effects of additive iron on growth,tissue distribution,haematology and immunology of gilthead sea bream,Sparus aurata[J].Aquacult Int,2010,18:1093-1104.
[20]ANDERSEN F,MAAGE A,JULSHAMN K.An estimation of dietary iron requirement of Atlantic salmon,Salmo salar L.,parr[J].Aquaculture Nutrition,1996,2:41-47.
[21]KAWATSU H.Studies on the anemia of fish:Ⅴ.dietary iron deficient anemia in brook trout,Salvelinus fontinalis[J].Bulletin of Freshwater Fish Research Laboratory,1972,22(1):59-67.
[22]SAKAMOTO S,YONE Y.Iron deficiency symptoms of carp[J].Bulletin of the Japanese Society of Scientific Fisheries,1978,44(10):1157-1160.
[23]SAKAMOTO S,YONE Y.Requirement of red sea bream for dietary iron-Ⅱ[J].Bulletin of the Japanese Society of Scientific Fisheries,1978,44(3):223-225.
[24]DAVIS D A,LAWRENCE A L.Evaluation of the dietary iron requirement of Penaeus vannamei[J].Journal of the World Aquaculture Society,1992,23(1):15-22.
[25]CHU J H,CHEN S M,HUANG C H.Effect of dietary iron concentrations on growth,hematological parameters,and lipid peroxidation of soft-shelled turtles,Pelodiscus sinensis[J].Aquaculture,2007,269:532-537.
[26]CHU J H,CHEN S M,HUANG C H.Growth,haematological parameters and tissue lipid peroxidation of soft-shelled turtles,Pelodiscus sinensis,fed diets supplemented with different levels of ferrous sulphate[J].Aquaculture Nutrition,2009,15(1):54-59.
[27]LIM C,SEALEY W M,KLESIUS P H.Iron methionine and iron sulfate as sources of dietary iron for channel catfish Lctalurus punctatus[J].Journal of the World Aquaculture Society,1996,27(3):290-296.
[28]LING J,FENG L,LIU Y,et al.Effect of dietary iron levels on growth,body composition and intestinal enzyme activities of juvenile Jian carp(Cyprinus carpio var.Jian)[J].Aquaculture Nutrition,2010,16(6):616-624.
[29]张佳明.鲈鱼(Lateolabrax japonicus)和大黄鱼(Pseudosciaena crocea R.)微量元素——锌、铁的营养生理研究[D].硕士学位论文.青岛:中国海洋大学,2007.
[30]乔永刚.军曹鱼微量元素锌、铁、铜营养生理的研究[D].硕士学位论文.青岛:中国海洋大学,2007.
[31]LIM C,KLESIUS P H,LIM H,et al.Interaction between dietary levels of iron and vitamin C on growth,hematology,immune response and resistance of channel catfish(Ictalurus punctatus)to Edwardsiella icta-luri challenge[J].Aquaculture,2000,185:313-327.
[32]VANGEN B,HEMRE G I.Dietary carbohydrate,iron and zinc interactions in Atlantic salmon(Salmo salar)[J].Aquaculture,2003,219:597-611.
[33]ANDERSEN F,LYGREN B,MAAGE A,et al.Interaction between two dietary levels of iron and two forms of ascorbic acid and the effect on growth,antioxidant status and some non-specific immune parameters in Atlantic salmon(Salmo salar)smolts[J].Aquaculture,1998,161:437-451.
[34]GAMBLING L,DANZEISEN R,FOSSET C,et al.I-ron and copper interactions in development and the effect on pregnancy outcome[J].The Journal of Nutrition,2003,133:1554S-1556S.
[35]SOLOMONS N W.Competitive interaction of iron and zinc in the diet:consequences for human nutrition[J].The Journal of Nutrition,1986,116:927-935.
[36]GUNSHIN H,MCKENZIE B,BERGER U V,et al.Cloning and characterization of a mammalian protoncoupled metal-ion transporter[J].Nature,1997,388:482-488.
[37]OGINO C,YANG G Y.Requirement of rainbow trout for dietary zinc[J].Bulletin of the Japanese Society of Scientific Fisheries,1978,44(9):1015-1018.
[38]GUDLAUG T,JAKOB K,SIGURLAUG S,et al.Copper,ceruloplasmin,superoxide dismutase and iron parameters in Parkinson’s disease[J].Pharmacology& Toxicology,1999,85:239-243.
[39]LANNO R P,SLINGER S J,HILTON J W.Maximum tolerable and toxicity levels of dietary copper in rainbow trout(Salmo gairdneri Richardson)[J].Aquaculture,1985,49:257-268.
[40]CONRAD M E,UMBREIT J N,MOORE E G,et al.Separate pathways for cellular uptake of ferric and ferrous iron[J].American Journal of Physiology-Gastrointestinal and Liver Physiology,2000,279:767-774.
[41]MARJA M,ANTTI S.Changes in plasma lysozyme and blood leucocyte levels of hatchery-reared Atlantic Salmon(Salmo salar L.)and sea trout(Salmo trutta L.)during parr-smolt transformation[J].Aquaculture,1992,106:75-78.
[42]HOEPELMAN I M,JAARSMA E Y,VERHOEF J,et al.Effect of iron on polymorphonuclear granulocyte phagocytic capacity:role of oxidation state and effect of ascorbic acid[J].British Journal of Haematology,1988,70(4):495-500.
[43]周立斌,王安利,马细兰,等.饲料中铁对美国红鱼(Sciaenops ocellatus)生长和免疫的影响[J].海洋与湖沼,2009,40(5):663-668.
[44]SEALEY W M,LIM C,KLESIUS P H.Influence of the dietary level of iron from iron methionin and iron sulfate on immune response and resistance of channel catfish to Edwardsiella ictaluri[J].Journal of the World Aquaculture Society,1997,28(2):142-149.
[45]WEINBERG E D.Iron depletion:a defense against intracellular infection and neoplasia[J].Life Sciences,1992,50:1289-1297.
[46]魏万权,李爱杰,李德尚.饲料中添加铁对牙鲆幼鱼生长的影响[J].水产学报,1999,23:100-103.
[47]YOO J H,MAENG H Y,SUN Y K,et al.Oxidative status in iron-deficiency anemia[J].Journal of Clinical Laboratory Analysis,2009,23(5):319-323.
[48]刘扬.维生案C与幼建鲤消化吸收能力及其抗氧化作用关系研究[D].博士学位论文.雅安:四川农业大学,2009.
猜你喜欢
中国饲料(2021年17期)2021-11-02
成都大学学报(自然科学版)(2021年1期)2021-05-22
养殖与饲料(2020年8期)2020-02-17
湖南饲料(2019年4期)2019-10-17
河北渔业(2017年9期)2017-10-16
中国饲料(2017年7期)2017-01-17
动物营养学报(2015年10期)2015-12-01
动物营养学报(2015年10期)2015-12-01
百科知识(2014年23期)2014-09-10
中国海洋大学学报(自然科学版)(2014年6期)2014-02-28
