
基于数值模拟的栓塞修复过程中AE信号分析
2012-09-18 01:27:26郝燕华张祥雪丁小康
东北林业大学学报 2012年4期
郝燕华 张祥雪 丁小康 刘 姣
(北京林业大学,北京,100083)
植物体内水分运输的主要通道是导管(或管胞),由于植物叶片的蒸腾作用使木质部水流常处于负压状态下,当木质部导管中水势下降并低于某一阈值时就会发生空穴化现象并导致导管栓塞。水分胁迫、土壤质地、土壤养分、冻融交替和机械损伤等都会导致植物栓塞[1-2],其中最常见的是水分胁迫[3]。栓塞对植物最直接的影响就是阻断水分在植物体内的运输,植物木质部导水率可下降20%,干旱诱发的栓塞可使植物根系水导率和气孔导度分别损失80%和45%[4],甚至会造成植物死亡。但只要栓塞化程度在一定限度内无论木本植物或草本植物,当水势升高时栓塞化的导管或管饱都能够修复[5]。
空穴化过程中伴随有超声发射(ultrasonic acoustic emission,UAE)现象。Milburn和 Johnson首次用改进的听筒听取了植物空穴化过程中伴随发生的声发射(acoustic emission,AE)现象[6]。这一发现实现了空穴化过程的实时监测,促进了栓塞研究的快速进展。近年来,学者们关于空穴化过程中植物超声发射的情况与水势、温度、光照、蒸腾速率等环境因子的关系进行了大量的研究[7-9]。但在研究空穴化超声发射的过程中意外地发现了在水势上升栓塞修复的过程中也可以监测到大量AE信号。Tyree在玉米的研究结果中发现,在每日15—18时水势升高期间,仍监测到了较多的AE信号[10],Raschi对茄子和Yang对西红柿的研究中也发现了类似的结果[11-12]。贾秀领等反复验证了一日内水势回升期小麦叶片上可测到相当数量的AE信号,且信号发生量和叶片水势回升有正相关趋势[7]。可以看出水势恢复期AE信号的发生并非偶然现象,长期以来研究者们一直关注的是空穴化过程中的AE信号,而这些水势回升期的信号高峰极可能与栓塞修复过程有关[7]。为了探究这个问题,笔者运用数值模拟的方法在Rayleigh-Plesset方程基础上建立描述栓塞修复过程中空泡的运动方程,对空穴化后的气泡进行动力学分析,进而探究栓塞修复过程中产生AE信号的原因。
1 空泡运动方程的建立
Zimmermann提出的“空气接种学说”(air seeding)认为,木质部管道栓塞是由来自外界大气空间或者在已栓塞化的木质部管道内的空气,经由管道间纹孔膜上的微孔充散至充水管道内所形成[13]。用Rayleigh-Plesset方程描述进入木质部液中空气种子在气泡内外压强作用下的半径变化[14]。为了满足Rayleigh-Plesset方程且简化计算,设空气种子在运动过程中保持球形对称,半径为r(t),其中t为时间。内部气体满足理想气体状态方程,气泡体积变化瞬间的空气溶解和蒸发可以忽略,气泡内外的温度梯度与热辐射对计算结果的影响较小可不考虑,则可认为运动过程为等温过程[14]。考虑室温下气泡的运动情况,设气泡半径为r0(r0为气泡初始半径)时的压强为PG0,即气泡的初始压强为PG0,则气泡内部压强PB(t)可以表示为[14]:
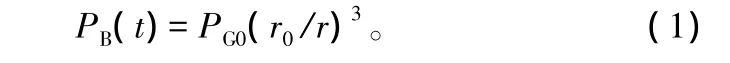
引入Rayleigh-Plesset方程,其中 P∞(t)可以看作等于气泡外木质部液的绝对压强PL。得到描述栓塞修复过程中空泡运动的最终方程:
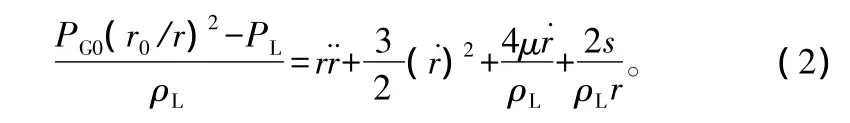
式中:ρL为导管中液体密度,近似等于水的密度;μ为动态黏滞系数,假设该值为常数且恒定;S为表面张力系数。
2 数值模拟结果
将常温293K下物性参数值:ρL=998 kg/m3,μ=1.002×10-3kg·m-1·s-1,S=0.073 N·m-1代入方程(2)。根据Zimmermann得出的结论,发生空穴化后气泡内压强为饱和蒸汽压[13],因此此时方程(2)中的PG0=0.0023 MPa。为计算方便认为空穴化后的气泡仍然保持球形对称。采用四阶龙格—库塔法求解空泡运动方程(2),可以具体描述在PL增大的过程中,即木质部液水势上升过程中,栓塞修复时空化泡的运动情况。
若发生空穴化后的气泡半径为30 μm,在木质部液为-100 Pa时,经方程(2)计算得到空化泡在修复过程中的半径变化。计算结果如图1所示,可以读出经一次塌缩后的半径为20.8 μm,气泡塌缩过程中其半径不是单调减小,而是以减幅振动方式达到新的平衡。气泡的减幅振动可以认为是一个振源,产生声发射事件。根据Zimmermann的理论,平衡后气泡内的一部分气体溶解于水,使气泡内部压强又回到饱和蒸汽压。此时假设木质部液压强升高到0,气泡会继续塌缩成半径为11.9 μm的气泡,如图2所示;压强继续升高为100 Pa,气泡继续塌缩,缩小到半径为5.16 μm的气泡,如图3所示;当压强继续升高到1000 Pa时,气泡会继续塌缩,变为更小的气泡,如图4所示,气泡半径为1.27 μm。以上每一次气泡塌缩都伴随声发射事件。
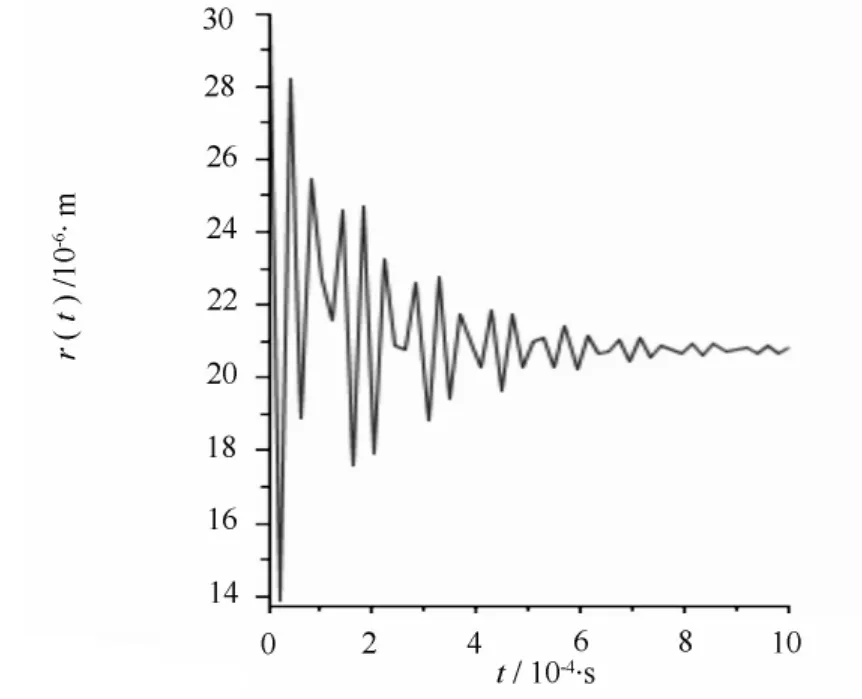
图1 当r=30 μm,pL=-100 Pa空化泡塌缩时的半径变化
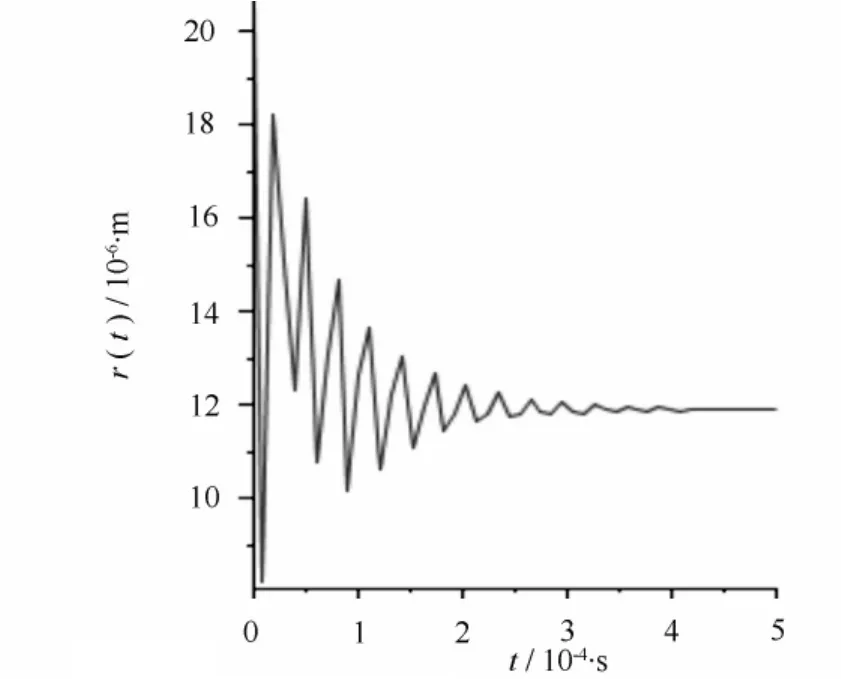
图2 当r=20.8 μm,pL=0空化泡塌缩时的半径变化
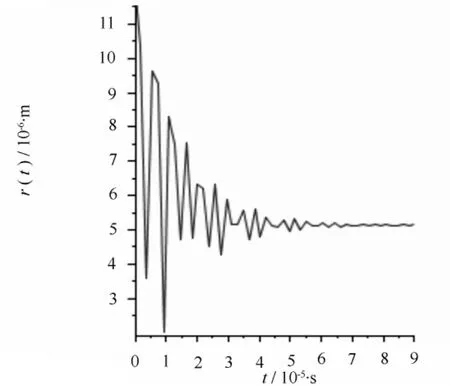
图3 当r=11.9 μm,pL=100 Pa空化泡塌缩时的半径变化
当木质部液水势为-100 Pa时,假设空化泡初始半径为40、50 μm。计算得到塌缩后的半径为32.1、45.1 μm,如图5、图6所示。可以看出初始半径不同的空化泡在相同压强下的修复情况不同。
为了验证数值模拟的结果,使用自组装的基于LabVIEW的超声发射检测系统在实验室检测植物木质部在栓塞修复过程中的AE信号,并将其与数值模拟结果对比。测量用的仪器是美国物理声学公司的声发射检测系统,传感器是该公司的WD型传感器。前置放大器是该公司的2/4/6C型放大器。软件部分基于LabVIEW做了扩展,实现的主要功能包括信号采集与保存、小波降噪、频谱分析、波形与频率分析等。
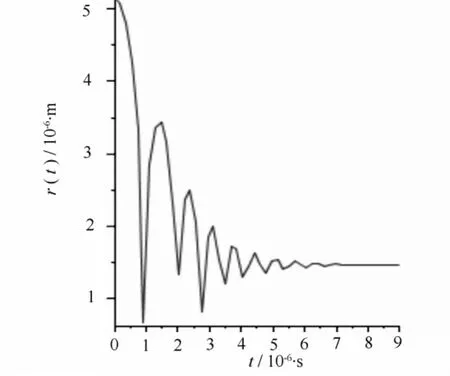
图4 当r=5.16 μm,pL=1000 Pa空化泡塌缩时的半径变化
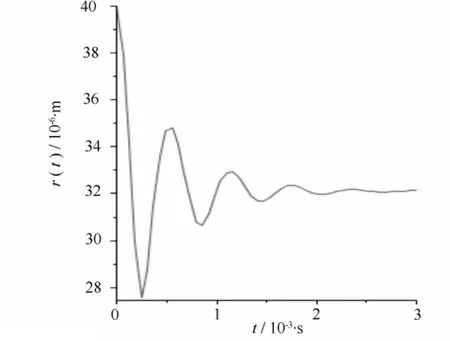
图5 当r=40 μm,pL=-100 Pa空化泡塌缩时的半径变化
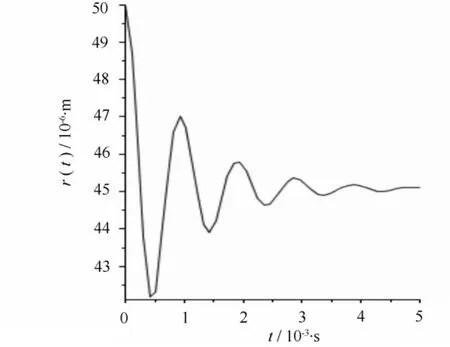
图6 当r=50 μm,pL=-100 Pa空化泡塌缩时的半径变化
实验材料采用夏季生长良好的侧柏(Platycladus orientalis(Linn.)Franco)、元宝枫(Acer truncatum Bunge)、刺槐(Robinia pseudoacacia L.)、栾树(Koelreuteria paniculata)。选取这4个树种阳面树冠下部直径为1.5 cm左右,长1.5 m左右的枝条,其中一部分用于离体枝条测量,另一部分用于切片测量。
比较离体枝条测量得到的数据和切片测量得到的数据发现两种方法得到的信号特征基本一致,但离体枝条测量耗时长,可重复性差且不易于进行定量研究。故为了进一步对AE信号进行研究,采用切片测量得到的数据进行分析。图7为检测到的4个树种切片栓塞修复过程中AE信号的典型波形图。
3 讨论
根据图1—图4的计算结果可以将栓塞修复过程看作是气泡在木质部液压强不断升高的过程中逐步塌缩减小的过程。如图1所示,空化泡经过一次塌缩过程后半径相对于导管来说仍然是比较大的,栓塞修复过程显然没有完成。当木质部液压强升高,空泡就会在前一次塌缩后的基础上继续塌缩,形成更小的气泡,如图2—图4所示。当木质部液压强升高时,空化泡经多次塌缩,半径不断减小,直到形成不影响水分输运的稳定的微小气泡,最终解除栓塞。
比较图1、图5和图6,在相同压强下,初始半径为30、40、50 μm的气泡经一次塌缩后半径分别减少了30%、19.75%、9.8%,Lewis等提到半径较小的气泡在相同压强下更容易修复[15],这个计算与该结论相符。但要想得到更确切的结论还需进一步的实验与计算论证。
比较图1—图6,可以看出栓塞在水势达到正压或稍小负压时就可以修复。而且在不同的压强和不同空化泡半径下,气泡塌缩时半径并不都是单调减小,而是以减幅运动的方式不断振动,这种振动会引起压强变化,从而形成振源,产生AE信号。声传感器将这种振动转化为电信号的变化,进而监测到AE信号。从计算结果可以看出,不同情况下得到的AE信号波形特征基本相似。
从7图得到,4个树种的实测声发射信号波形基本相似,振铃计数均在10个左右,且信号均以减幅振动的形式衰减,这与用数值模拟得到的波形图(图1—图6)特征是一致的。
4 结论
植物木质部栓塞修复过程监测到的AE信号,其来源尚不清楚,也未见文章讨论过。本研究通过对空化泡动力学方程进行计算,认为可将植物木质部栓塞修复过程看作是空穴化后的气泡在木质部液压强不断升高过程中逐步塌缩减小的过程。空泡塌缩的过程中其半径是以减幅振动的方式减小。气泡的减幅振动可以形成振源,产生AE信号。此信号是栓塞修复过程的AE信号来源之一。计算结果还表明栓塞在水势达到正压或稍小负压时就可以修复。气泡越小,越容易修复。

图7 4个树种木质部栓塞修复过程中AE信号的典型波形图
[1]Meinzer F C,Clearwater M J,Goldstein G.Water transport in trees:current perspectives,new insight and some controversies[J].Environmental and Experimental Botany,2001,45:239-262.
[2]Mayr S,Hacke U,Schmid P,et al.Forst drought in conifers at the alpine timberline xylem dysfunction and adaptations[J].Ecology,2006,87(12):3175-3185.
[3]Crombie D S,Milbum J A,Hipkins M F.Maximum sustainable xylem sap tensions in Rhododendron and other speices[J].Planta,1985,163:27-33.
[4]李卫民,张佳宝.植物木质部导管栓塞[J].植物生理学通讯,2008,44(3):581.
[5]孙青.几种木本植物木质部栓塞修复机制的进一步研究[D].北京:北京林业大学,2006:14-15.
[6]Tyree M T,Sperry J S.Vulnerability of xylem to cavitation and embolism[J].Annual Review Plant Physiology and Plant Molecular Biology,1989,40:19-38.
[7]贾秀领,马瑞竟,张全国,等.冬小麦叶片气栓塞与叶水势日变化的关系[J].麦类作物学报,2005,25(1):50-54.
[8]霍晓静,钱东平,吕长飞,等.用声发射技术实现作物生理需水信息监测[J].节水灌溉,2008(9):10-13.
[9]Zweifel R,Zeugin F.Ultrasonic acoustic emissions in drought-stressed trees-more than signals from cavitation[J].New Phytologist,2008,179:1070-1079.
[10]Tyree M T,Fiscus E L,Wullschleger S D,et al.Detection of xylem cavitation in corn under field conditions[J].Plant Physiology,1986,82:597-599.
[11]Yang S F.Development of an automatic and real-time watering system based on estimation of water stress in tomato crops with acoustics emission technology[J].Transaction of the CSAE,2001,20:40-50.
[12]Raschi A,Torri F,Zienna P,et al.Cavitation events in Solanum melongena L.measured after water stress cycles in field conditions[J].Acta Horticulturea,1990,278:119-128.
[13]Tyree M T,Zimmermann M H.Xylem structure and the ascent of sap[M].Berlin:Springer-Verlag,2002:44-47.
[14]Christopher E Brennen.Cavitation and bubble dynamics[M].New York:Oxford University Press,1995:47-50.
[15]Lewis A M,Harnden V D,Tyree M T.Collapse of water-stress emboli in the tracheids of Thuja occidentalis L[J].Plant Physiology,1994,106(4):1639-1646.
猜你喜欢
钢铁钒钛(2023年5期)2023-11-17 08:48:34
植物研究(2021年2期)2021-02-26 08:40:10
厦门理工学院学报(2016年1期)2016-12-01 04:50:53
水利科技与经济(2016年7期)2016-04-25 13:03:00
水利科技与经济(2016年3期)2016-04-09 13:57:20
水利科技与经济(2016年4期)2016-04-09 03:49:18
水利科技与经济(2016年2期)2016-04-09 02:09:13
天津农业科学(2015年11期)2015-12-03 00:03:49
江苏农业科学(2015年4期)2015-06-15 21:11:35
安徽农学通报(2014年7期)2014-04-29 03:00:24
