
不同生境黄鼬毛皮保温性能比较及传热测试实验优化1)
2012-09-18 01:27:26张溯源
东北林业大学学报 2012年4期
张溯源 张 伟 郑 雷
(东北林业大学,哈尔滨,150040)
热导率是反映其传递热量能力大小的综合指标,热导率越大,保温性能越弱[1]。目前,国际上已经有相关学者利用自制的实验装置对动物毛被的热导率、传热热阻、导热热阻、冷却系数、均温系数等指标进行测定[2-6]。生物体与环境之间物质和能量的交换过程是生命系统最基本的过程之一;在这一过程中,热能必须经过动物内环境与外界环境的临界,即动物的体被[7-8]。同种生物在不同分布地域、性别、季节、身体部位方面,其被毛性状具有差异,并在阻止机体热量向外界散失上起到了积极的作用[9-14]。
在以往的实验过程中,有若干问题对实验结果的准确性造成影响,如毛皮样本的干硬皱褶,在实验室难以取得平整的实验试样,使对同一样本进行多次重复操作测得的热导率方差较大。另外,实验测试的持续时间多长可以准确表现样本的热导率,保证标定冰点的状态便于测试数据的准确等都需要进行传热测试实验的优化,使实验误差尽可能减小,以得到准确的动物毛皮传热能力的量化信息。为此,本文在动物体被稳态过程热物性参数测试平台构建完成的基础上[15],以通河林区与哈尔滨平原地区的黄鼬冬季雄性皮张为实验材料,开展两种环境下生长的小型哺乳动物毛被热导率的比较研究,试图以热导率说明不同环境下的同一物种在毛皮保温性能上的差异。
1 研究地气候概况
通河林区位于哈尔滨平原北部,在北纬44°~46°之间。林区年温差较大,最大温差可达83.6℃;全年平均气温1.9℃,1月极端最低气温为-46.2℃。最大积雪深度是1977年12月30日的38 cm。10月下旬到翌年5月下旬为结冻期,长达7个月。冻土深度1.6~1.9 m,最深1.93 m。哈尔滨平原平均气温为2.3℃;1月份为全年最冷月,平均气温-21.1℃,极端最低气温-42.6 ℃;平均年温差 42.9℃;冷冻期为5个月。平地最大积雪深度33 cm;冻土深度 1.8 ~2.0 m[16-17]。
2 材料与方法
随机选取11—12月份哈尔滨平原、黑龙江通河林区的雄性成体黄鼬毛皮各10张。
于黄鼬生皮上取100 mm长、50 mm宽的试样,利用动物体被稳态过程热物性参数测试平台进行传热测试。通过测试平台倒出的EXCEL表格,计算每采集间隔内所测试冷热面温差与热流密度测试平均值,带入公式K(热导率)=1/((冷热面温差/热流密度)+1/23),计算热导率。每个试样测试3次,取平均值作为最终热导率。
3 结果与分析
3.1 测试实验条件的优化
实验毛皮样本的采样时间、身体部位和性别选择:分布在黑龙江地区的黄鼬在9月上旬开始脱夏毛长冬毛,冬毛的稳定持续期为10月下旬到翌年3月。选择11—12月份这一时间段的黄鼬皮张作为实验样本,既可以保证动物冬季毛被达到稳态,又可以防止被毛因长期执行保护保温功能带来的损伤所造成的影响。动物背臀部毛被对于机体保温占有重要地位,黄鼬的冬毛先从臀部长出,其次是背部和两胁,即背臀部先于身体其他部位换上冬毛[19]。所以,本实验选取黄鼬的背臀部毛皮作为实验样本。在平整的整张生皮的尾基部上方30 mm处,以体轴为对称轴,制取宽50 mm、长100 mm的背臀部毛皮作为热导率测定样本。本实验全部选择雄性个体的黄鼬,一方面保证样本的可比性;另一方面因雄性个体较大,更有利于保障实验样本的面积。
测试样本的预处理:因采集的生皮干硬且不平整,难以防止试样与均热板之间进入空气,而空气是热的不良导体,对实验结果的准确性影响较大[20]。所以,利用0.9%的生理盐水浸泡采集的黄鼬生皮,进行24 h回鲜处理。将回鲜后的毛皮展平(避免拉伸)并固定在毛皮展架上,自然干燥72 h并避免阳光直射。自然风干后即为平整的生皮,可用于热导率测试样本的取样。0.9%的生理盐水接近哺乳动物正常组织液浓度,毛皮在进行回鲜过程中不会因为细胞吸水而发生膨胀;并且避免了在自然干燥过程后,细胞失水较多造成毛皮收缩现象[21]。整个预处理过程对毛皮无物理和化学伤害。
测试持续时间的确定:为了确定测试样本的最适时间,分别随机选取通河林区、哈尔滨平原地区的黄鼬冬季毛皮样本各1张,进行10 h的热导率测定,数据采集间隔为300 s;然后代入公式计算每2 h的热导率均值进行比较,从而确定每次实验测试的持续时间。由图1、图2可以看出,当测试时间为120 min后,两个样本通过毛被试样的热量交换已进入稳态。因此,略去对初始波动较大2 h的热导率均值计算,计算剩余4组2 h热导率均值进行比较发现;所得结果相对偏差小于1%。所以,当进行毛被热导率对比时,测试持续时间为4 h即可。

图1 通河林区随机黄鼬个体10 h数据
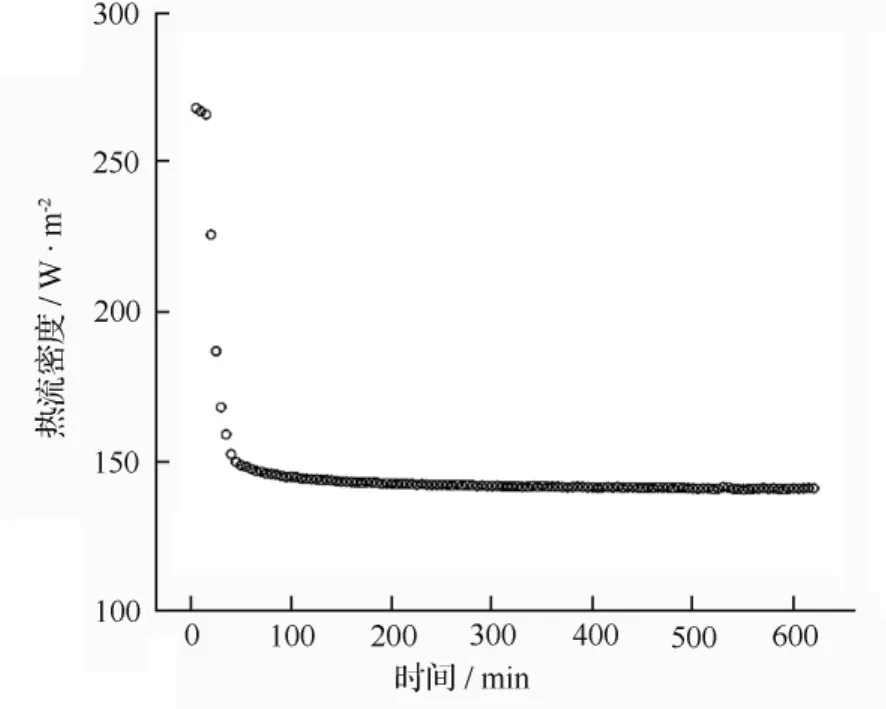
图2 哈尔滨平原随机黄鼬个体10 h数据
实验冰点的标定:用于标定的冰点一定为冰水混合物,实验中需要保证冰点状态。若冰水混合物中的冰溶尽,则会造成热流密度取值较正常冰点的取值偏大,影响准确性。对同一试样分别进行有冰点和无冰点的热流密度测试(见图3、图4),有冰点的测试数值在80 W/m2浮动,无冰点的同一试样测试数值在100 W/m2浮动,差异显著。

图3 有冰点测试数据
传热测试实验优化后的条件:在整个传热测试实验过程中,确定了用0.9%生理盐水对样本进行24 h回鲜和72 h的自然风干预处理;选取雄性黄鼬背臀部5000 mm2面积的毛皮进行传热性能测试,每次测试持续时间为4 h;标定冰点的状态为冰水混合物。

图4 无冰点测试数据
3.2 保温性能比较
3.2.1 不同生境冬季毛皮热导率差异
利用动物体被稳态过程热物性参数测试平台进行测试,每个样本测试3次,每次测试时间为4 h。对通河林区、哈尔滨平原20只黄鼬冬季毛皮样本的热导率测试结果见表1。

表1 通河林区与哈尔滨平原黄鼬毛皮的热导率
对表1热导率利用SPSS17.0软件进行独立样本 T 检验[18],结果显示:P 值为 0.002,小于显著性水平0.05,故拒绝零假设,认为两总体均值存在显著差异,即通河林区的黄鼬在毛皮传热性能(平均热导率3.92 W·m-1·K-1)上要明显低于哈尔滨平原地区(平均热导率 4.35 W·m-1·K-1)。
3.2.2 同一地区不同采样时间段的冬季毛皮热导率差异
为检验同一地区不同采样时间的样本的传热性能是否存在显著差异,按采样时间以11月15日为分界,将来自哈尔滨平原的10个样本分为组1、组2;以11月和12月为分界,将来自通河林区的10个样本分为组3、组4;每组均为5个样本。分别比较组1和组2、组3和组4,传热性能的差异。分组见表2。
由于是小样本量,采用非参数检验中的两独立样本曼-惠特尼U检验组1和组2、组和、组4,检验两独立样本是否来自于无显著差异的总体。结果表明:组1和组2的U统计量的精确概率P值为0.421,大于显著差异值,故两独立样本来自于无显著差异的总体,即来自哈尔滨平原的2组毛皮样本在热导率上差异不显著。组3和组4的U统计量的精确概率P值为0.310,大于显著差异值,也表明来自通河林区的2组毛皮样本在热导率上差异不显著。
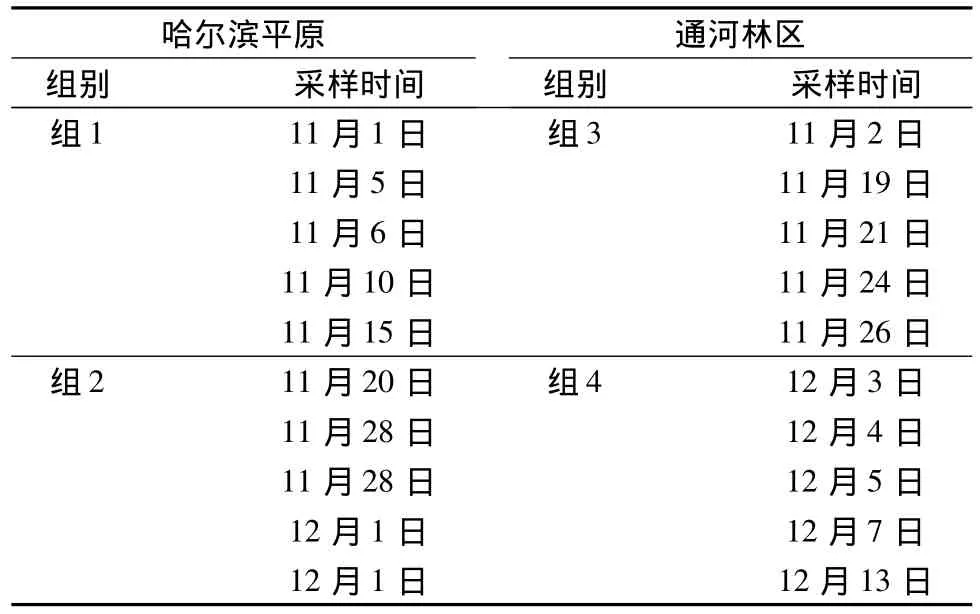
表2 相同地区不同采样时间段的样本分组
4 结论与讨论
黄鼬东北亚种在2种生境条件下的雄性成体冬季毛皮热导率均值差异显著,且来自通河林区的保温性能优于来自哈尔滨平原。对于来自同一地区不同采样时间段的2组毛皮样本的热导率比较显示,11—12月份的同一地区的冬季毛皮保温性能差异不显著,即冬季毛被已于10月末进入稳态。同一亚种黄鼬在山区和平原2种不同生境下,其毛被在形态学上差异比较显著,如密度、细度等指标[22]。本实验所获得的量化的毛皮热导率,鲜明地反映了毛被形态学差异所带来的保温功能差异。通河林区的黄鼬东北亚种的冬季毛皮要比哈尔滨平原地区的保温能力强,是2种生境的黄鼬种群分别适应不同环境条件的需要。两地没有明显的纬度和海拔高度差异,主要是生态环境的不同;通河林区进入冬季时间早,秋季气温下降快,整个冬季平均气温低,且持续时间比平原地区长。正是这些环境差异,使得同一亚种黄鼬的不同种群出现了毛皮性状上的不同,以满足不同的保温功能需要。
本实验通过对不同地区毛皮导热率的量化处理以及对实验数据的分析,比较出不同地区毛被形态学差异所带来的保温功能差异。在实验中,通过对实验过程的优化处理,减小了因实验操作而产生的误差,从而减小对毛皮导热率量化处理的影响,使数据更精确。
[1]陶文铨.传热学[M].西安:西北工业大学出版社,2006.
[2]Scholander P F,Walters V,Hock R,et al.Body insulation of some arctic and tropical mammals and birds[J].Biol Bull,1950,99:225-236.
[3]vadsheim P H K,Folkow L P,Blix A S.A new device for measurement of the thermal conductivity of fur and bulbber[J].Journal of Thermal Biology,1994,19(6):431-435.
[4]Carolyn J G,Caroline M,Onischak,et al.Acute early thermal experience alters weaning onset in rats[J].Physiology & Behavior,1998,64(4):463-474.
[5]Justin G B,George S B.Seasonal changes and wind dependence of thermal conductance in dorsal fur from two small mammal species(Peromyscus leucopus and Microtus pennsylvanicus)[J].Journal of Thermal Biology,2007,32:383-387.
[6]Robyn S H,Brenda A D,Linda G F,et al.Body temperature,thermoregulatory behaviour and pelt characteristics of three colour morphs of springbok[J].Comparative Biochemistry and Physiology-Part A:Molecular& Integrative Physiology,2009,152(3):379-388.
[7]刘静,王存诚.生物传热学[M].北京:科学出版社,1997.
[8]张伟,景松岩,徐艳春.毛皮学[M].哈尔滨:东北林业大学出版社,2002.
[9]周宏力.龙江路、辽宁路元皮的质量差异[J].东北林业大学学报,2004,32(3):55-58.
[10]杨晓东,任露泉.动物毛发的形态结构及其功能特性研究[J].农业工程学报,2002,18(2):21-24.
[11]张伟,孙长虹,马跃,等.通河林区黄鼬背中部与爪部冬季上毛的形态结构[J].兽类学报,2008,28(4):440-444.
[12]华彦,张伟,徐艳春.黑龙江小兴安岭地区黄鼬夏冬季毛皮性状比较[J].兽类学报,2010,30(1):110-114.
[13]程志斌,张伟,华彦,等.漠河地区养殖的北极狐冬季被毛性状与保温性能的关系[J].生态学报,2010,30(11):2972-2980.
[14]郑雷,张伟,华彦.哺乳动物毛被传热性能及其影响因素[J].生态学报,2011,31(13):3856-3862.
[15]郑雷,张伟,张斌,等.动物体被稳态过程热物性参数测试平台构建[J].东北林业大学学报,2011,39(2):117-119.
[16]葛亮.黄鼬东北亚种三个主要分布地的毛皮性状与生态因子的比较分析[D].哈尔滨:东北林业大学,2008.
[17]刘玉莲,周海龙,苍蕴琪.四季分明是哈尔滨气候的显著特征[J].黑龙江气象,2003(3):21-28.
[18]张红兵,贾来喜,李潞.SPSS宝典[M].北京:电子工业出版社,2007:170-236.
[19]朱婧,王家义.黄鼬换毛序[J].动物学杂志,1960(7):293-295.
[20]杨世铭.传热学[M].北京:高等教育出版社,1987:7-32.
[21]林加涵,魏文铃,彭宣宪.现代生物学实验:上册[M].北京:高等教育出版社,2000:70-87.
[22]金煜,张梦利,李少白,等.黄鼬东北亚种山皮甸皮差异原因分析[J].野生动物,1996(3):15-17.
猜你喜欢
储能科学与技术(2022年5期)2022-05-10 10:18:50
黑龙江大学自然科学学报(2022年1期)2022-03-29 00:57:48
装备制造技术(2020年2期)2020-12-14 03:09:12
陶瓷学报(2020年5期)2020-11-09 09:23:04
活力(2019年15期)2019-09-25 07:21:08
福建稻麦科技(2018年2期)2018-03-31 00:22:51
儿童时代·幸福宝宝(2018年12期)2018-01-21 17:33:20
农村百事通(2016年22期)2016-12-17 19:39:44
农村百事通(2016年18期)2016-11-05 16:35:27
农村百事通(2015年24期)2015-12-28 11:11:33
