
二氯化钴致人结肠癌细胞系CacyBP/SIP核转位
2012-09-15 08:00谢福利仇长青赵盈盈冯珊珊翟惠虹
基础医学与临床 2012年11期
谢福利,仇长青,赵盈盈,杨 博,冯珊珊,翟惠虹*
(1.宁夏师范学院医学院,宁夏固原756000;2.宁夏医科大学临床学院总医院消化内科,宁夏银川750004)
低氧在结肠癌发生过程中具有重要的作用[1],可促进结肠癌细胞增殖、抑制凋亡及增强化疗耐药[2],但具体机制不清。CacyBP/SIP自1996年发现以来,越来越多的证据显示[3]:在肿瘤发生、发展、侵袭及转移等方面都有重要的作用;同时,具有依赖Ca2+浓度的核转位现象[4],核转位后促进结肠癌细胞增殖[5]。低氧是否可诱导CacyBP/SIP核转位?本研究拟通过构建CacyBP/SIP过表达慢病毒颗粒,转染至结肠癌细胞,给予 CoCl2刺激,观察CacyBP/SIP在结肠癌细胞中的定位。
1 材料与方法
1.1 材料
结肠癌细胞SW480(上海中科院);CoCl2(Sigma公司);胞质胞核蛋白抽提试剂盒(Pierce公司);CacyBP/SIP mAb(Cell signaling公司);293T细胞、pGC-LV重组载体、pHelper 1.0和pHelper 2.0(吉凯基因公司);其他相关试剂(Fermentas、SinoBio、Takara、NEB、捷瑞公司)。
1.2 方法
1.2.1 过表达CacyBP/SIP慢病毒载体构建:应用人结肠癌细胞系HCT-116的cDNA序列,采用Primer Premier v5.0软件设计上游和下游引物,用PCR法扩增CacyBP/SIPcDNA序列。带有绿色荧光的慢病毒载体与目的基因分别用NheⅠ和EcoRⅠ进行双酶切,T4连接酶将酶切后的产物相连,连接产物转化大肠杆菌DH5α感受态细胞,应用PCR法筛选阳性克隆,接种阳性转化子,送测序。用 Lipofectamine2000共转染pGC-LV载体和两种辅助包装原件载体至293T细胞包装。培养8h更换为完全培养基,培养48 h后,收集上清液,浓缩后在293T细胞中采用逐孔稀释法测定并标定病毒滴度。
1.2.2 转染结肠癌细胞:常规培养传代,在T50瓶细胞长至30% ~50%时,每瓶加入500 μL(5 mg/L)Polybrene、4 400 μL完全培养基和100 μL(1 ×108个细胞)过表达CacyBP/SIP的慢病毒颗粒,放入培养箱培养8~12 h弃去细胞上清,更换为新鲜培养基继续培养。至细胞长至80%以上汇合时,正常消化传代并行激光共聚焦检测。
1.2.3 激光共聚焦检测:取3个12孔培养板,每孔放置一个细胞爬片,每孔接种(3~4)×104个细胞。至细胞长至最佳状态时,吸干培养基,用PBS冲洗2次,分别加入2 mL含 50、100 及150 μmol/L CoCl2培养基。对照组每孔分别加入完全培养基作为对照。分别在培养箱培养0.5和1 h。用4%多聚甲醛1 mL,固定20 min,PBS冲洗3遍,再用0.1%的 Tritont 100,37℃温箱内打孔45 min,PBS冲洗3遍,最后用含DAPI封片剂染色封片。激光共聚焦检测CacyBP/SIP在细胞中的定位。
1.2.4 Western blot法检测:对于激光共聚焦检测阳性结果(100 μmol/L CoCl2刺激1 h),胰蛋白酶消化后以500 r/min离心3 min,取细胞沉淀用胞质蛋白、核蛋白抽提试剂盒分别抽提胞质、胞核蛋白,用BCA试剂盒进行蛋白定量;上样缓冲液稀释至5 g/L、煮沸后 -80℃冻存;把20 μL蛋白样品上样到12%SDS-PAGE胶加样孔内,电泳电压80 V,时间20 min;湿式转膜,电流200 mA,60 min;加入5%脱脂牛奶封闭60 min后,加入 CacyBP/SIP单抗(1∶200)4℃孵育过夜,常规洗膜后加入羊抗鼠IgG(1∶2 000)的二抗,室温孵育1 h,洗涤。曝光、显影。
2 结果
2.1 构建了过表达CacyBP/SIP慢病毒载体
PCR扩增目的基因(图1);对构建的过表达CacyBP/SIP慢病毒载体阳性克隆进行PCR鉴定(图2),与目标基因测序比对(图3);绘制熔解曲线(图4)。


2.2 转染结肠癌SW480细胞
将过表达CacyBP/SIP的慢病毒颗粒,转染至结肠癌SW480细胞,复感染指数(MOI)为20,96 h后激光共聚焦观察到绿色荧光(图5)。
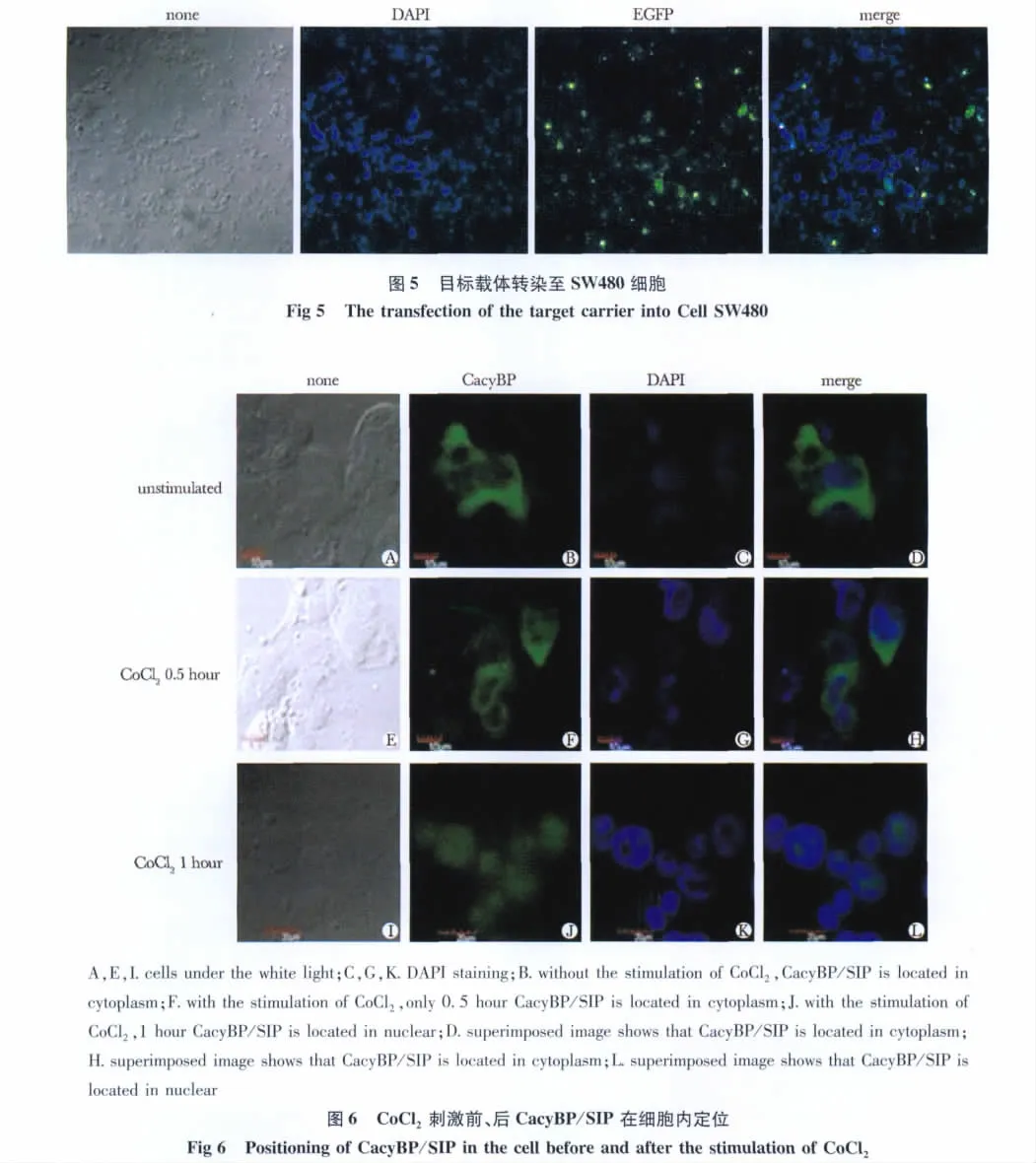
2.3 激光共聚焦检测CoCl2刺激前、后CacyBP/SIP在细胞内的定位
无CoCl2刺激条件下 CacyBP/SIP主要位于细胞胞质内;给予100 μmol/L CoCl2刺激0.5 h,CacyBP/SIP位于胞质;刺激1 h时观察到CacyBP/SIP转位至胞核(图6)。
2.4 Western blot检测CoCl2刺激前、后CacyBP/SIP在细胞内的定位
无刺激条件下,CacyBP/SIP主要在胞质内表达,给予100 μmol/L CoCl2刺激1 h,可在细胞核内表达(图7)。
3 讨论

图7 CoCl2刺激前、后 Western blot检测CacyBP/SIP在胞质及胞核中的表达Fig 7 The expressions of CacyBP/SIP in cytoplasm and nuclear under the detecting by Western blot before and after the stimulation of CoCl2
CacyBP/SIP分子质量约26 ku,含一个687 bp的开放阅读框,编码228个氨基酸,存在7个磷酸化位点和1个核定位信号[6]。其功能主要与红细胞的发育[7]和新的泛素-蛋白酶体降解有关[8]。与肿瘤的关系以往研究发现,CacyBP/SIP在胃癌阿霉素耐药细胞中高表达并参与胃癌细胞多药耐药,其下调可以部分逆转耐药性[9]。近年来又发现CacyBP/SIP参与乳腺癌、胰腺癌的发生发展及复发转移,可能成为预后不良的生物学指标之一[10-11]。因此推断CacyBP/SIP与恶性肿瘤的发生发展密切相关。
在前期研究中利用淋巴细胞杂交瘤技术制备了3株CacyBP/SIP单克隆抗体,对正常及肿瘤组织中表达研究发现[12]:CacyBP/SIP在结肠癌组织中的表达明显高于癌旁组织,在正常结肠组织不表达,且主要定位于胞质及胞核。
CacyBP/SIP具有依赖 Ca2+浓度的核转位现象[13]。那么,究竟哪些因素可以引起 CacyBP/SIP核转位?
人体肿瘤存在低氧的假设从上世纪50年代年提出,80年代得到证实。肿瘤中的低氧表现为不稳定性。后多家研究证实低氧在结肠癌发生过程中都具有重要作用[14]:低氧可促进结肠癌细胞增殖、凋亡抑制、增强化疗耐药、迁移能力增强。Co2+是铁螯合酶的底物,可替代氧感受器血红素中的Fe2+,在高浓度时与氧结合,将此分子锁定在脱氧合状态,使细胞在不缺氧的环境下“感觉”缺氧。因此本研究用CoCl2刺激来模拟结肠癌低氧,观察低氧状态下CacyBP/SIP是否发生核转位?
本研究通过CacyBP/SIP过表达慢病毒载体转染至结肠癌SW480细胞,激光共聚焦显微镜发现CacyBP/SIP主要位于胞质,Western blot亦证实其位于胞质。给予不同浓度CoCl2,人为制造细胞低氧刺激,发现低氧时,CacyBP/SIP可发生核转位现象,Western blot亦证实CacyBP/SIP在刺激前位于胞质,刺激后部份位于胞核。
蛋白在细胞中的定位常常与其功能密切相关,很多蛋白分子从无活性形式变为活性形式时,会发生分子定位的变化,进入到特定的亚细胞结构中发挥其功能。CacyBP/SIP在给予刺激后发生核转位及磷酸化,提示其可能做为信号分子传递信息,从而参与细胞功能的调节。前期已发现CacyBP/SIP在结肠癌中高表达,免疫组化结果提示CacyBP/SIP位于胞浆及胞核,低氧可诱导CacyBP/SIP核转位,CacyBP/SIP是否做为信使,传递上游促进恶性肿瘤增殖的信号,需进一步研究。
[1]Jiang YA,Fan LF,Jiang CQ,et al.Expression and significance of PTEN,hypoxia-inducible factor-1 alpha in colorectal adenoma and adeno-carcinoma[J].Word J Gastroenterol,2003,9:491 -494.
[2]刘祺,龚凡杰,谭泽明,等.低氧诱导对人结肠癌SW480细胞低氧诱导因子-1α和Survivin基因表达的影响[J].中华实验外科杂志,2006,23:1228 -1230.
[3]ZHai H,Shi Y,Jin H,et al.Expression of calcyclin-bindingprotein/Siah-1 interacting protein in normal and malignant human tissues:an immunohistochemical survey[J].J Histochem Cytochem,2008,56:765 -772.
[4]Filipek A,Jastrzebska B,Nowotny M,et al.Ca2+-dependent translocation of the calcyclin-binding-protein in neurons and neuroblastoma NB-2a cells[J].J Biol Chem,2002,277:21103-21109.
[5]翟惠虹,陈雄,卢媛媛,等.结肠癌细胞中CacyBP/SIP的表达及核转位现象[J].世界华人消化杂志,2008,16:3953-3957.
[6]Filipek A,Kuznicki J.Molecular cloning and expression of a mouse brain cDNA encoding a novel protein target of cal-cyclin[J].J Neuroehem,1998,70:1793-1798.
[7] Pircher TJ,Geiger JN,Zhang D,et al.Wojchowski DM.Integrative signaling by minimal erythropoietin receptor forms and c-Kit[J].J Biol Chem,2001,276:8995 -9002.
[8] Matsuzawa SS,Takayama BA,Froesch JM,et al.Reed.P53-inducible human homologue of Drosophila seven in absentia(Siah)inhibits cell growth:suppression by BAG-1[J].EMBO J,1998,17:2736 -2747.
[9]Yongquan Shi,Wenhua Hu,Fang Yin,et al.Regulation of drug sensitivity of gastric cancer cells by human calcyclinbinding-protein(CacyBP)[J].Gastric Cancer,2004,7:160-166.
[10]王宁菊,马刚,翟惠虹.钙周期素结合蛋白与乳腺癌临床的相关性研究[J].第四军医大学学报,2009,30:2403-2407.
[11]Wang N,Ma Q,Wang Y,et al.CacyBP/SIP expression is involved in the clinical progression of breast cancer[J].World J Surg,2010,34:2545-52.
[12]Chen X,Mo P,Li X,et al.CacyBP/SIP protein promotes proliferation and G1/S transition of human pancreatic cancer cells[J].Mol Carcinog,2011,50:804 -810.
[13]Zhai H,Shi Y,Jin H et al.Expression of calcyclin-binding-protein/Siah-1-interacting-protein in normal and malignant human tissues:an immunohisto-chemical survey[J].J Histochem Cytochem,2008,56:765 -772.
[14]Jing Wu,Xinyu Tan,Xiaozhong Peng,et al.Translocation and phosphorylation of calcyclin binding protein during retinoic acid-induced neuronal differentiation of neuroblastoma SH-SY5Y cells[J].J Biochemi Mol Biol,2003,36:354-358.
猜你喜欢
天津医科大学学报(2021年3期)2021-07-21
西南国防医药(2016年7期)2016-12-01
天津医药(2016年9期)2016-10-20
国外医药(抗生素分册)(2016年5期)2016-07-12
中国卫生标准管理(2015年1期)2016-01-14
中国老年学杂志(2015年16期)2015-03-05
癌变·畸变·突变(2015年4期)2015-02-27
实验动物与比较医学(2014年5期)2014-02-28
河南医学研究(2014年3期)2014-02-27
植物营养与肥料学报(2011年4期)2011-10-26
