
高致病性猪繁殖与呼吸综合征病毒强弱毒ORF1a、ORF1b、ORF2-ORF7片段互换嵌合病毒的构建及其生物学特性的分析
2012-09-11 07:22:16姜一峰周艳君王亚欣朱建平徐彦召虞凌雪童光志
中国预防兽医学报 2012年1期
姜一峰,周艳君,王亚欣,朱建平,徐彦召,童 武,虞凌雪,童光志
(中国农业科学院上海兽医研究所猪病研究室,上海200241)
猪繁殖与呼吸综合征病毒(Porcine reproductive and respiratory virus,PRRSV)属于尼多病毒目,动脉炎病毒科,动脉炎病毒属,属单股正链RNA病毒[1]。基因组全长约15kb,5'端有帽子结构,3'端有polyA尾,病毒非结构蛋白编码区靠近5'端,占基因组全长约4/5,病毒结构蛋白编码区靠近3'端,部分结构蛋白基因首尾有部分重叠[2]。
PRRSV有9个开放阅读框(ORF),分别为ORF1a、 ORF1b、 ORF2、 ORF2a、 ORF3、 ORF4、ORF5、 ORF6、 ORF7, 分 别 编 码 pp1ab、 pp1a、GP2、E、GP3、GP4、GP5、M、N蛋白[1-3]。最近的研究显示,在ORF4与ORF5上存在一个新的ORF,命名为ORF5a,编码新发现的结构蛋白GP5a[4-5]。ORF1a,ORF1b编码病毒的非结构蛋白,ORF2-7编码病毒结构蛋白。
2006 年,我国猪群中出现了一种变异的PRRSV,与经典的PRRSV相比毒力显著增强[6],将该高致病性PRRSV(HP-PRRSV)在体外多次传代后毒力显著下降,而且对猪不致病[7]。本实验以HP-PRRSV强毒HuN4-F5及其体外传代致弱病毒HuN4-F112的感染性分子克隆为基础,将ORF1a,ORF1b,ORF2-73个大片段在强弱毒之间进行相互替换,以研究ORF1a,ORF1b,ORF2-7对病毒生物学特性的影响。
1 材料和方法
1.1 病毒株、细胞及主要试剂 HP-PRRSV HuN4-F5株及其弱毒HuN4-F112株感染性分子克隆由本实验室构建[8-9];BHK-21、Marc-145细胞由本实验室保存;羊抗鼠FITC荧光标记的IgG(FITC-TgG)购自北京中杉金桥生物技术有限公司;转染试剂DMRIE-CReagent购自Invitrogen;DMEM、Opti-MEM培养基购自GIBCO公司;体外转录试剂盒mMessage High Yield Capped RNA Transcription kit购自Ambion公司;PRRSV阳性血清由本实验室制备保存;DL2000DNA Marker购自TaKaRa公司。
1.2 嵌合病毒的构建策略 HuN4-F5和HuN4-F112全病毒基因组分别被克隆在质粒pBluescriptⅡSK(+)中,选择PacⅠ(位于质粒上)和AscⅠ酶切位点作为强弱毒ORF1a互换所用酶切位点,NheⅠ和AscⅠ作为强弱毒ORF1b互换所用酶切位点,AscⅠ和NotⅠ(位于质粒上)作为强弱毒ORF2-7互换所用酶切位点(图 1)。

1.3 嵌合病毒基因组的体外转录 将PRRSV全基因组连接到质粒pBluescriptⅡ SK(+)时在3'末端引入了SwaⅠ酶切位点,在转染之前,将包含嵌合病毒全长基因组的质粒用SwaⅠ酶切线性化。体外转录按试剂盒操作说明进行,转录反应体系为:10μL 2×NTP/CAP,2μL Enzyme M ix,2μL 10×Reaction buffer,1μg线性化质粒模板,加无核酸酶水至20μL,置于37℃水浴锅中转录1h 45min。
1.4 嵌合病毒的拯救 按照转染试剂DMRIE-C Reagent操作说明方法,将转录产物转染细胞密度达到70%~90%的BHK-21细胞。37℃,5%CO2培养4h,吸出上清液,再加入2%~3%FBS的细胞维持液。24h后收取细胞,-80℃保存。
取转染后冻存的BHK-21细胞冻融离心后,上清接入6孔板内Marc-145细胞单层中,置CO2培养箱内孵育1h~1.5h后换含2%~3%FBS的细胞维持液。每5d为一代次进行拯救病毒传代培养,传5代直到细胞病变(CPE)出现为止。
1.5 拯救病毒的鉴定 本实验室在构建PRRSV HuN4-F5株和HuN4-F112株的感染性克隆时,将基因组第14680位的A沉默突变为G产生一个M luⅠ酶切位点作为鉴定拯救病毒的分子标记。根据该酶切位点两侧的序列设计了一对PCR扩增引物,参照文献[8]和[9]方法鉴定。
1.6 间接免疫荧光试验(IFA) 将拯救的病毒接种于Marc-145细胞单层中,培养48h后,用80%乙醇固定30m in,以PRRSV阳性血清(1∶100稀释)为一抗,37℃作用1h,以羊抗鼠FITC-IgG为二抗,37℃避光作用1h,PBS洗涤后荧光显微镜下观察。
1.7 重组病毒生长曲线的绘制 将拯救的重组病毒以100TCID50感染M arc-145细胞,分别在感染后12h、24h、36h、48h、60h、72h、84h 和 96h收取 300μL细胞上清液,使用 100μL测定其TCID50,计算病毒滴度,根据不同时间点病毒的滴度绘制病毒生长曲线。TCID50的测定方法是将病毒液做连续10倍稀释(10-1~10-10),然后接种于96孔培养板培养的MARC-145细胞单层上。每个稀释度接种8孔,即做8个重复。同时设两组阴性对照。在37℃5%CO2条件下培养,记录CPE情况。结果依据Reed-Muench法计算TCID50。
2 结果
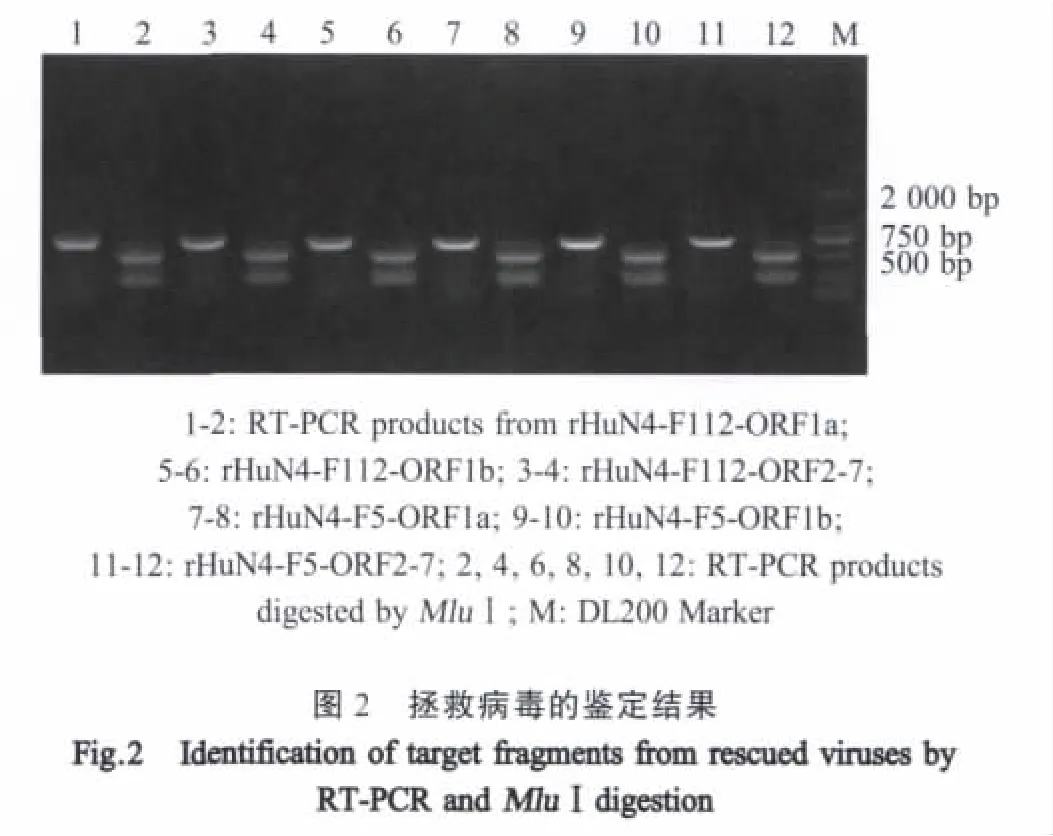
2.1 PRRSV强弱毒嵌合病毒的拯救及鉴定 以HP-PRRSV HuN4-F5及其弱毒HuN4-F112的感染性分子克隆为骨架,将ORF1a、ORF1b和ORF2-73个基因片段相互替换,形成的6种嵌合病毒基因组,以强毒株为骨架的命名为HuN4-F5-ORF1a,HuN4-F5-ORF1b和HuN4-F5-ORF2-7;以弱毒株为骨架的分别命名为HuN4-F112-ORF1a,HuN4-F112-ORF1b和HuN4-F112-ORF2-7。将这些嵌合病毒基因组先转染BHK细胞,再在Marc-145细胞上传代,在5代内均出现CPE,表明已拯救出病毒。提取拯救病毒核酸,经RT-PCR扩增,M luⅠ酶切鉴定,结果出现两条带,表明获得的病毒为感染性分子克隆拯救获得的嵌合病毒(图2)。部分测序结果显示,ORF1a、ORF1b、ORF2-7都得到了正确的嵌合。
采用PRRSV阳性血清对拯救的重组病毒rHuN4-F5-ORF1a、 rHuN4-F5-ORF1b、 rHuN4-F5-ORF2-7、 rHuN4-F112-ORF1a、 rHuN4-F112-ORF1b、rHuN4-F112-ORF2-7进行IFA检测,拯救的嵌合病毒均能够被PRRSV阳性血清识别,出现特异性荧光反应(图 3)。

2.2 PRRSV强弱毒嵌合病毒在细胞上的生长特性将拯救的病毒按10倍稀释接种生长Marc-145细胞的96孔组织培养板,观察CPE,并按Reed-Muench法计算病毒滴度(表1)。以强毒株为骨架的重组病毒在Marc-145上的生长滴度都低于相应的以弱毒株为骨架的重组病毒。

表1 拯救的重组PRRSV在细胞上的生长滴度Table 1Replication titer of the rescued PRRSVs
测定各拯救病毒接种Marc-145细胞后96h内的生长曲线,结果显示,以强毒株为骨架的重组病毒除rHuN4-F5-ORF1a生长滴度显著升高以外,另外两个替换(rHuN4-F5-ORF1b和 rHuN4-F5-ORF2-7)与亲本病毒(rHuN4-F5)相比没有明显变化(图4A)。而以弱毒株为骨架的重组病毒除rHuN4-F112-ORF1b与亲本病毒(rHuN4-F112)相比没有明显变化以外,另外两个替换的重组病毒(rHuN4-F112-ORF2-7和rHuN4-F112-ORF1a)生长滴度均有显著下降(图4B)。
3 讨论
PRRSV基因组全长约15kb,非结构蛋白编码区约占基因组全长的4/5,另外1/5为结构蛋白编码区。非结构蛋白又可分为 ORF1a、ORF1b。在ORF1a中包含PRRSV两个重要的酶,nsp2和nsp4,他们负责切割多聚蛋白pp1ab和pp1a,形成14个非结构蛋白,这些非结构蛋白在PRRSV的复制中起重要作用[1,10,12]。Nsp1、nsp2可以抑制宿主干扰素的产生,调节机体免疫系统[11-13]。Nsp1进入细胞核参与了PRRSV各个复制阶段的串联作用[10-12]。Nsp2作为最大的非结构蛋白,在免疫调节以及病毒的复制以及病毒毒力变化上都起着重要作用[10,14-15]。Nsp3-8可能是的毒力基因[16]。ORF1b中包含RNA病毒复制起始的两个重要蛋白RdRP及解旋酶,他们组成了PRRSV复制起始的转录翻译复合体[17-18]。Nsp11作为内切核酸酶,需要严格的时间、位置限定,以避免切割病毒本身RNA,可能是潜在的自杀基因[10,19]。ORF2-7编码病毒的结构蛋白,在病毒的侵入细胞、进入细胞核、病毒释放以及免疫逃避中起重要作用[2,20]。
本实验分别以PRRSV HuN4强毒株及其传代致弱病毒HuN4F112株在感染猪时,强毒是致死性的,弱毒不致病;而在体外细胞培养时弱毒的增殖滴度明显高于强毒。比较强弱毒基因组序列时发现,弱毒株共有77个氨基酸发生突变,其中41个发生在ORF1a区,15个发生在ORF1b区,21个发生在ORF2-7。强弱毒的差异正是由于这77个氨基酸的突变决定的,因此本实验设计分别以强弱毒的感染性分子克隆为骨架,将这3个大片段在强弱毒之间互换,期望发现哪个功能区的变化将影响PRRSV在细胞中的适应性,进而影响病毒的致病性。结果发现以强毒株为骨架替换弱毒株的ORF1a后,在Marc-145中其增殖滴度显著提高,而以弱毒株为骨架替换强毒株的ORF1a后,在细胞中的增殖滴度显著下降,这表明ORF1a区与PRRSV在Marc-145细胞中的适应性密切相关。本实验结果将为进一步在体内研究ORF1a、ORF1b、ORF2-7的变化对PRRSV毒力的影响奠定了基础。
[1]Snijd er E J,Meulenberg J J M.The molecular biology of arteriviruses[J].JGen Virol,1998,79:961-979.
[2]Dea S,Gagnon C A,Mardassi H,et al.Current know ledge on the structural proteins of porcine reproductive and respiratory syndrome(PRRS)virus:comparison of the North American and European isolates[J].Arch Virol,2000,145:659-688.
[3]Wu Wai-hong,Fang Ying,Farwell R,et al.A 10-kDa structural protein of porcine reproductive and respiratory syndrome virus encoded by ORF2b[J].Virology,2001,287:183-191.
[4]Firth A E,Zevenhoven-Dobbe JC,W ills N M,et al.Discovery of a small arterivirus gene that overlaps the GP5coding sequence and is important for virus production[J].J Gen Virol,2011,92:1097-1106.
[5]Johnson C R,Griggs T F,Gnanandarajah J,et al.Novel structural protein in porcine reproductive and respiratory syndrome virus encoded by an alternative ORF5present in all arteriviruses[J].JGen Virol,2011,92:1107-1116.
[6]Tong Guang-zhi,Zhou Yan-jun,Hao Xiao-fang,et al.Highly pathogenic porcine reproductive and respiratory syndrome,China[J].Emerg Infect Dis,2007,13(9):1434-1436.
[7]Tian Zhi-jun,An Tong-qing,Zhou Yan-jun,et al.An attenuated live vaccine based on highly pathogenic porcine reproductive and respiratory syndrome virus(HP-PRRSV)protects piglets against HP-PRRS[J].Vet M icrobiol,2009,138:34-40.
[8]张善瑞,周艳君,朱建平,等.高致病性猪繁殖与呼吸综合征病毒HuN4株感染性分子克隆的建立及拯救病毒的鉴定[J].中国预防兽医学报,2011,33(7):497-502.
[9]Zhang Shan-rui,Zhou Yan-jun,Jiang Yi-feng,et al.Generation of an infectious clone of HuN4-F112,an attenuated live vaccine strain of porcine reproductive and respiratory syndrome virus[J].Virol J,2011,8:410-416.
[10]Fang Ying,Snijder E J.The PRRSV replicase:Exploring the multifunctionality of an intriguing set of nonstructural proteins[J].Virus Res,2010,154:61-76.
[11]Kroese M V,Zevenhoven-Dobbe JC,Bos-de Ruijter JN,et al.The nsp1alpha and nsp1papain-like autoproteinases are essential for porcine reproductive and respiratory syndrome virus RNA synthesis[J].JGen Virol,2008,89:494-499.
[12]Tijms M A,Snijder E J.Equine arteritis virus nonstructural protein 1,an essential factor for viral subgenom ic mRNA synthesis,interacts w ith the cellular transcription cofactor p100[J].JGen Virol,2003,84:2317-2322.
[13]Beura L K,Sarkar S N,Kwon B,et al.Porcine reproductive and respiratory syndrome virus nonstructural protein 1betamodulates host innate immune response by antagonizing IRF3activation[J].Virology,2010,84:1574-1584.
[14]Chen Zhen-hai,Zhou Xiao-xin,Lunney JK,et al.Immunodominant epitopes in nsp2of porcine reproductive and respiratory syndrome virus are dispensable for replication,but play an important role in modulation of the host immune response[J].J Gen Virol,2010,91:1047-1057.
[15]Faaberg K S,Kehrli Jr M E,Lager K M,et al.In vivo grow th of porcine reproductive and respiratory syndrome virus engineered nsp2deletion mutants[J].Virus Res,2010,154:77-85.
[16]Kwon B,Ansari IH,Pattnaik A K,et al.Identification of virulence determ inants of porcine reproductive and respiratory syndrome virus through construction of chimeric clones[J].Virology,2008,380:371-378.
[17]O'Reilly E K,Cheng Kao C.Analysis of RNA-dependent RNA polymerase structure and function as guided by known polymerase structures and computer predictions of secondary structure[J].Virology,1998,252:287-303.
[18]Bautista E M,Faaberg K S,M ickelson D,et al.Functional properties of the predicted helicase of porcine reproductive and respiratory syndrome virus[J].Virology,2002,298:258-270.
[19]Nedialkova D D,Ulferts R,Van Den Born E,et al.Biochem ical characterization of arterivirus nonstructural protein 11reveals the nidovirus-wide conservation of a replicative endoribonuclease[J].Virology,2009,83(11):5671-5682.
[20]Das P B,Dinh P X,Ansari IH,et al.The m inor envelope glycoproteins GP2a and GP4of porcine reproductive and respi-ratory syndrome virus interact w ith the receptor CD163[J].Virology,2010,84:1731-1740.
猜你喜欢
昆明医科大学学报(2022年2期)2022-03-29 00:51:20
科学大观园(2022年2期)2022-01-23 11:05:15
华侨大学学报(自然科学版)(2021年4期)2021-07-30 02:18:58
现代仪器与医疗(2021年2期)2021-07-21 02:19:16
中国保健营养(2021年16期)2021-04-03 18:15:56
中国当代医药(2020年26期)2020-11-06 07:22:10
生物信息学(2020年2期)2020-07-09 01:27:56
动物医学进展(2015年10期)2015-12-07 05:46:18
分析化学(2015年10期)2015-11-03 07:38:01
现代检验医学杂志(2015年2期)2015-02-06 02:01:10
