
HIF-1α基因沉默对大鼠肝癌CBRH-7919细胞p27和Ki67表达的影响*
2012-08-02 13:04许林锋倪嘉延陈耀庭孙宏亮吴裕丹
中国病理生理杂志 2012年10期
许林锋,倪嘉延,陈耀庭,孙宏亮,吴裕丹
(中山大学孙逸仙纪念医院 1介入放射科,2血液科,广东 广州 510120)
缺氧诱导因子 1α(hypoxia inducible factor 1α,HIF-1α)是由 Semenza等[1]于1992 年在缺氧诱导的细胞核抽提物中发现的一种重要的核转录因子。HIF-1由HIF-1α和HIF-1β 2个亚基组成,HIF-1α决定HIF-1的活性。HIF-1α在恶性肿瘤信号转导通路中扮演重要的角色,是肿瘤细胞对缺氧适应反应的中心环节,与肿瘤的侵袭和转移密切相关。p27蛋白为细胞周期的重要负性调控因子,可最直接地影响检查点的调控,进而调控细胞的增殖活性[2]。Ki67蛋白是一种与细胞周期相关的增殖细胞核杭原,Ki67标记指数能可靠地反映正常和病变组织的增殖活性,对肿瘤的复发和转移有预测作用[3]。本研究探讨大鼠肝癌CBRH-7919细胞在缺氧环境下,HIF-1α基因沉默对p27和Ki67表达及细胞增殖的影响。
材料和方法
1 材料
大鼠肝癌细胞株CBRH-7919由中山大学实验动物中心提供。RPMI-1640培养基购自Gibco;脂质体LipofectamineTM2000购自Invitrogen;NC膜购自Pall;HIF-1α抗体购自Santa Cruz;p27和Ki67抗体购自CST;β-actin抗体购自博士德公司;TRIzol试剂购自TaKaRa;RT-PCR试剂盒购自TaKaRa;HIF-1α和β-actin引物用Primer 3软件设计,由Invitrogen公司合成。
2 方法
2.1 细胞培养 CBRH-7919细胞用含10%胎牛血清的RPMI-1640培养液,在37℃、5%CO2、饱和湿度培养箱连续培养、传代。将生长状态良好的肝癌细胞消化下来以(8~10)×104/L密度接种于6孔板于37℃、5%CO2、饱和湿度培养箱中培养24 h后,加入150 μmol/L CoCl2溶液,用于模拟肿瘤内部缺氧环境。
2.2 HIF-1α siRNA载体的构建 根据前期实验研究,选择GenBank中大鼠HIF-1α基因编码区序列作为分析序列。采用RNAdraw分析软件对序列的二级结构进行预测,然后按照Elbashir等推荐的siRNA设计原则进行设计,并筛选出一条HIF-1α靶核苷酸序列(靶mRNA)。P1:5’-GAAACUCUUCCAAGCAAUUtt- 3’,P2:5’ - AAUUGCUUGGAAGAGUUUCtt-3’,参照siRNA的基本设计原理选择靶序列,针对目的基因的序列设计互补的siRNA表达载体核苷酸链,即HIF-1α siRNA特异性干扰序列。将siRNA溶液与脂质体LipofectamineTM2000载体试剂混合,形成siRNA/LipofectamineTM2000载体复合物。
2.3 细胞转染 6孔板种植细胞(8~10)×104/L,在含10%胎牛血清的RPMI-1640培养液中培养24 h,细胞汇合度达到30% ~50%。将稀释好的HIF-1α siRNA和脂质体LipofectamineTM2000载体试剂混合,形成 HIF-1α siRNA/LipofectaminTM2000复合物。将HIF-1α siRNA转染载体加到含有细胞和培养基的培养板的孔中融合,然后放入37℃、CO2培养箱培养。6 h后换含10%血清的RPMI-1640培养液,20 h恢复生长后,荧光显微镜下观察细胞绿色荧光蛋白的表达以检测转染效率。转染分组:(1)空白对照组:仅加入LipofectamineTM2000试剂。(2)转染组:转染HIF-1α siRNA序列入细胞。
2.4 Real-time RT-PCR检测 TRIzol法提取6孔板内各组细胞的总RNA并定量;各样本取2 ng~2 μg总RNA,按TaKaRa公司试剂盒说明进行逆转录合成cDNA。应用real-time RT-PCR检测,βactin作为内参照。PCR引物,HIF-1α正义链ACTGCACAGGCCACATTCAT,反义链CGAGGCTGTGTCGACTGAGA;β-actin正义链 CCTAGGCACCAGGGTGTGAT,β-actin反义链 TTGGTGACAATGCCGTGTTC。反应体系为20 μL总体系,反应条件95℃ 3 min预变性,95℃ 10 s,55℃ 30 s,39个循环,并在每个循环延伸末端点收集荧光信号,绘制扩增曲线。
2.5 Western blotting检测 冷PBS洗涤细胞2次,加入全蛋白裂解液后冰上裂解10 min收获蛋白。将抽提蛋白用BCA法定量后,于100 V泳道进行SDS-PAGE;电泳结束后,以30 mA、120 min将蛋白转移至NC膜;封闭液(5%BSA/TBST)封闭60 min,加入Ⅰ抗(1∶3000稀释)4℃过夜,Ⅱ抗(1∶6000稀释)室温下杂交120 min,TBST洗膜3次后用ECL试剂盒进行发光反应,压片、显影、定影,观察蛋白印迹,并进行图像分析。
2.6 流式细胞术检测细胞周期 选择对数生长期细胞,用不含EDTA的胰蛋白酶收集处理后的细胞,800 r/min离心10 min,沉淀采用300 μL PBS重悬,逐滴加入700 μL预冷的无水乙醇中,乙醇终浓度为70%,4℃避光固定过夜。800×g离心10 min,去上清。PBS洗2次。重悬细胞于500 μL含1×105U/L RNase的PBS缓冲液中,避光,37℃孵育30 min。加2 g/L PI至终浓度50 mg/L,避光孵育30 min。流式细胞仪激发波长488 nm,发射波长525 nm检测。
3 统计学处理
应用SPSS 17.0统计分析软件。计量资料用均数±标准差()表示,两组间采用t检验,多个不同处理组组内、组间计量资料的比较应用单因素方差分析。以P<0.05为差异有统计学意义。
结 果
1 缺氧条件下肝癌细胞HIF-1α的表达
CBRH-7919细胞经150 μmol/L CoCl2处理12 h后,HIF-1α mRNA表达量较常氧培养组显著增高(P<0.05);24 h后细胞HIF-1α mRNA相对表达量进一步增加(P<0.05),见图1。CBRH-7919细胞HIF-1α蛋白的表达也呈显类似规律,见图2。
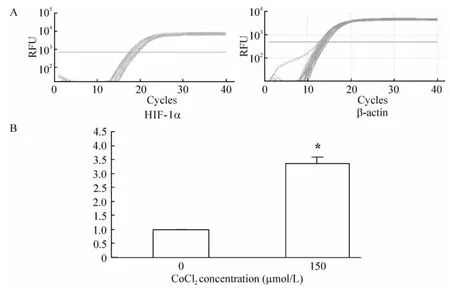
Figure1.Expression HIF-1α mRNA in hepatoma cells 24 h after treatment with CoCl2.A:real-time RT-PCR amplification curves of HIF-1α and β-actin under hypoxia;B:the change of HIF-1α mRNA expression in hepatoma cells..n=3.*P <0.05 vs 0 μmol/L.图1 肝癌细胞培养24 h后HIF-1α mRNA的表达
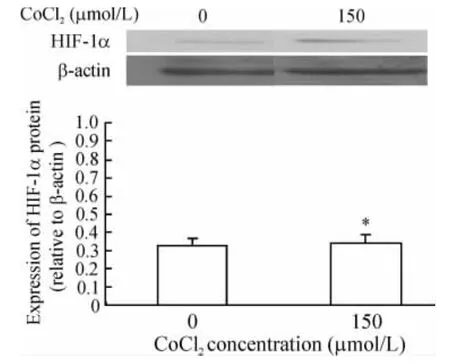
Figure2.Expression of HIF-1α protein in hepatoma cells treated with CoCl2for 24 h..n=3.*P<0.05 vs 0 μmol/L.图2 肝癌细胞HIF-1α蛋白的表达
2 siRNA干扰对HIF-1α表达的影响
在150 μmol/L CoCl2作用下,转染HIF -1α siRNA表达载体后HIF-1α mRNA表达明显减少,见图3,同步检测显示,HIF-1α蛋白表达亦受到明显抑制,见图4,与对照组比差异均具有统计学意义(P<0.05)。
3 沉默HIF-1α对p27表达的影响
150 μmol/L CoCl2处理和转染 HIF -1α siRNA表达载体后,p27蛋白表达较未转染HIF-1α siRNA表达载体的对照组明显增多,差异有统计学意义(P<0.05),见图4。

Figure3.The change of the mRNA expression of HIF-1α after HIF-1α siRNA transfection.A:real-time RTPCR amplification curves of HIF -1α;B:the mRNA expression of HIF-1α..n=3.*P<0.05 vs control.图3 HIF-1α siRNA干扰后肝癌细胞HIF-1α mRNA表达的变化
4 沉默HIF-1α对Ki67表达的影响
150 μmol/L CoCl2处理和转染 HIF -1α siRNA表达载体后,Ki67蛋白表达较未转染HIF-1α siRNA表达载体的对照组显著减少,两组差异有统计学意义(P<0.05),见图4。
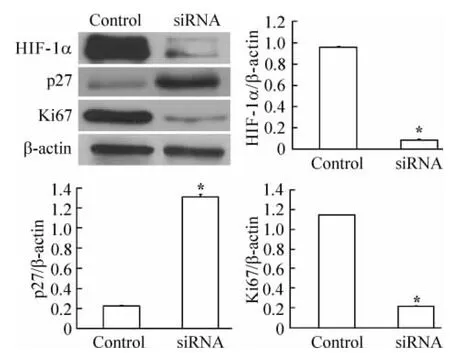
Figure4.The protein expression of HIF -1α,p27 and Ki67 after the transfection of HIF-1α siRNA..n=3.*P <0.05 vs control.图4 HIF-1α siRNA干扰后HIF-1α、p27和Ki67蛋白表达的变化
5 沉默HIF-1α对肝癌细胞增殖的影响
流式细胞术结果显示,HIF-1α siRNA转染组各期细胞分布为:G0/G1期68.7%,S期17.5%,G2/M期13.8%,而未转染siRNA表达载体的对照组各期细胞分布为:G0/G1期51.4%,S期32.7%,G2/M期15.9%。这表明HIF-1α siRNA能有效阻滞细胞于G0/G1期,进入S期的细胞数量明显减少。统计学分析显示,转染组与对照组的G0/G1期和S期细胞比例差异有统计学意义(均P<0.05),见图5。
讨 论
恶性肿瘤组织内部长期处于相对缺氧的微环境,可诱导HIF-1α的表达增加、降解减少。HIF-1α是肿瘤细胞对缺氧适应反应的中心环节,可上调多种靶基因的表达,使肿瘤对相对缺氧的微环境发生适应性改变。HIF-1α极不稳定,易被蛋白酶降解,常氧条件下半衰期较短,但在缺氧时显著延长[4]。
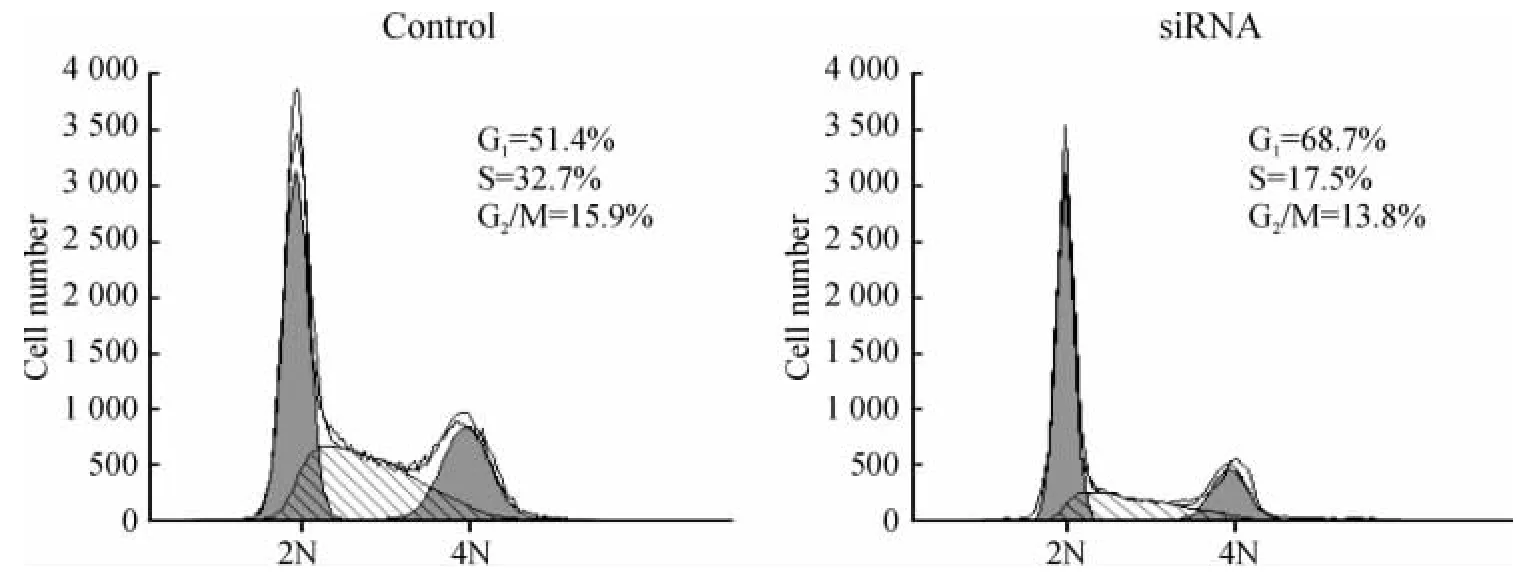
Figure5.Effect of HIF -1α silencing on cell cycle of hepatoma cells detected by flow cytometry.图5 流式细胞术检测HIF-1α siRNA干扰后肝癌细胞细胞周期的变化
根据以往学者相关研究经验,应用siRNA干扰技术,转染特异性siRNA于肿瘤细胞,可以有效沉默细胞中靶基因的表达。鉴于HIF-1α在恶性肿瘤细胞中的关键作用,本研究利用CoCl2建立缺氧模型,模拟肿瘤细胞内部相对缺氧的微环境,并转染特异性HIF-1α siRNA于大鼠肝癌CBRH-7919细胞。结果显示,缺氧环境下HIF-1α的表达显著增加;特异性siRNA干扰肿瘤细胞后,HIF-1α和Ki67的表达明显减少,p27的表达显著增加。这表明沉默HIF-1α在显著抑制Ki67表达的同时,明显促进了p27的表达。于此同时,肝癌细胞的增殖亦受到一定程度的抑制。
RNA干扰可以形象地称为基因沉默,是一种进化上保守的抵御转基因或外来病毒侵犯的防御机制,属于转录后基因沉默机制范畴[5],能特异性地封闭一个或多个基因,而不影响正常等位基因的功能,具有较高的选择性和特异性,减少非特异作用引起的副作用,是极富有前途的治疗手段。目前采用特异性siRNA沉默HIF-1α,探讨肝癌细胞中增殖相关因子表达变化的文献报道尚不多见。本研究发现HIF-1α基因沉默后,增殖相关因子p27蛋白的表达明显增多的同时Ki67蛋白的表达却显著减少。p27为细胞周期的重要负性调控因子,可最直接地影响限制点的调控,进而调控细胞的增殖活性。相关研究显示p27的表达量和肝细胞肝癌的恶性程度及复发转移密切相关[6-7]。Ki67是一种与细胞周期相关的增殖细胞核杭原,其标记指数能可靠的反映正常和病变组织的增殖活性,与肝细胞肝癌的病理特征及预后密切相关[8]。因此,沉默HIF-1α可能通过改变增殖相关因子的表达,使肿瘤细胞的增殖状态亦发生改变。
HIF-1α在血管内皮生长因子(vascular endothelial growth factor,VEGF)表达的信号转导通路中亦起着关键的作用。相关研究表明,HIF-1α在多个层次调节 VEGF的表达,其主要功能包括增强VEGF的转录活性与增加VEGF mRNA稳定性两个方面[9]。VEGF是最早被发现的可被缺氧环境诱导生成并促进肿瘤新生血管生成的生长因子[10-11]。HIF-1α通过上调VEGF的表达,诱导肿瘤新生血管的生成,与肿瘤的复发、转移密切相关。由此可见,利用siRNA干扰技术沉默肿瘤细胞HIF-1α的表达,可能对控制肿瘤的复发、转移起到重要的作用。
本实验研究尚存在着一定的局限性。应用特异性siRNA干扰技术沉默靶基因的表达,已有不少成功案例;但是截至目前为止siRNA干扰技术依然应用于离体实验阶段,在体动物实验或临床应用中,特异性siRNA干扰、沉默靶基因的功效如何,以及特异性siRNA载体的恰当选用,仍需要进一步的探索与证实。
[1]Semenza GL,Wang GL.A nuclear factor induced by hypoxia via de novo protein synthesis binds to the human erythropoietin gene enhancer at a site required for transcriptional activation[J].Mol Cell Biol,1992,12(12):5447-5454.
[2]Tesio M,Trumpp A.Breaking the cell cycle of HSCs by p57 and friends[J].Cell Stem Cell,2011,9(3):187 -192.
[3]Swarts DR,Ramaekers FC,Speel EJ.Molecular and cellular biology of neuroendocrine lung tumors:Evidence for separate biological entities[J].Biochim Biophys Acta,2012,1826(12):255 -271.
[4]Goteri G,Lucarini G,Montik N,et al.Expression of vascular endothelial growth factor(VEGF),hypoxia inducible factor-1alpha(HIF-1alpha),and microvessel density in endometrial tissue in women with adenomyosis[J].Int J Gynecol Pathol,2009,28(2):157 -163.
[5]Ingelbrecht I,Van Houdt H,Depicker A.Posttranscriptional silencing of report transgenes in tobacco correlates with DNA methylation[J].Proc Nail Acad Sci U S A,1994,91(22):10502 -10506.
[6]Tannapfel A,Grund D,Katalinic A,et al.Decreased expression of p27 protein is associated with advanced tumor stage in hepatocellular carcinoma[J].Int J Cancer,2000,89(4):350-355.
[7]邵 馨,顾春燕.P27和skp2在肝细胞癌中的表达及其意义[J].临床肝胆病杂志,2009,25(2):120 -121.
[8]Ebelt J,Neid M,Tannapfel A,et al.prognostiec significance of proliferation markers in hepatocellular carcinoma(HCC)[J].Zentralbl Chir,2000,125(7):597 -601.
[9]Ahluwalia A,Tarnawski AS.Critical role of hypoxia sensor:HIF-1α in VEGF gene activation.Implications for angiogenesis and tissue injury healing[J].Curr Med Chem,2012,19(1):90 -97.
[10]Feng J,Zhang Y,Xing D.Low -power laser irradiation(LPLI)promotes VEGF expression and vascular endothelial cell proliferation through the activation of ERK/Sp1 pathway[J].Cell Signal,2012,24(6):1116 - 1125.
[11]娄远蕾,刘 芬,高法梁,等.miR-210对血管内皮细胞VEGF-Notch信号通路的调控[J].中国病理生理杂志,2011,27(5):923 -927.
猜你喜欢
现代仪器与医疗(2022年4期)2022-10-08
华人时刊(2022年9期)2022-09-06
中国生殖健康(2020年2期)2021-01-18
华人时刊(2020年15期)2020-12-14
中国生殖健康(2018年2期)2018-01-12
中成药(2017年9期)2017-12-19
中成药(2017年7期)2017-11-22
中华老年多器官疾病杂志(2016年7期)2016-04-28
中国康复理论与实践(2015年10期)2015-12-24
中国火炬(2013年11期)2013-07-25
