
灰楸的繁育系统1)
2012-06-13 06:20李平英王军辉
东北林业大学学报 2012年2期
李平英 王军辉
(甘肃省小陇山林业科学研究所,天水,741022)(国家林业局林木培育重点实验室(中国林业科学研究院林业研究所))
韩云花 董菊兰 马丽娜 尚千涵 陈 静 于永明
(甘肃省小陇山林业科学研究所)
灰楸(Catalpa fargesiiBur)是紫葳科(Bignoniaceae)梓树属(Catatpa Scop)植物,自然分布于华北、西北和西南等地[1]。西北是灰楸的主要自然分布区,在甘肃省天水、平凉、陇南地区广泛分布。灰楸喜光,稍耐阴,速生。国内对灰楸的研究相对较少。对植物花结构和繁育系统的了解是认识植物生活史的前提,也是其他相关研究所必须依赖的知识背景。植物的繁育系统是代表所有直接影响后代遗传组成的有性特征的总和,主要包括花综合特征、花各性器官的寿命、花开放式样、自交亲和程度和交配系统,它们结合传粉者和传粉行为影响着生殖后代遗传组成和适合度,其中交配系统是核心[2]。植物繁育系统是当今进化生物学研究中最为活跃的领域。达尔文和最早的群体遗传学理论,都认为繁育系统对生物的遗传多样性和进化起重要作用。国内外学者对许多植物繁育系统进行了研究[5-12],对灰楸花的发育生物学特性研究未见报道。鉴于此,本研究对灰楸开花习性进行数年定位观测,以便准确掌握灰楸花期生物学特性,不失时机地收集花粉和适时进行人工授粉,为开展灰楸杂交育种提供理论指导。
1 试验地概况
2008、2009年在甘肃省小陇山林业科学研究所进行试验。海拔高度1 100 m,平均气温12℃,极端最高气温39℃,极端最低气温-19.2℃,相对湿度78%,年平均降水量700 mm,无霜期210 d;土壤为黄棕壤;属暖温带半湿润大陆性季风气候,光照充足,雨量偏少,6—8月份较干旱。试验对象为野生状态居群。所选树高8~12 m,地径16~35 cm,为开花结实盛期的成年植株。
2 材料与方法
2.1 花形态特征的观测
选择5~6株进行花的特性调查。选择开花植株中部花枝,用高枝剪剪下,观察、记录植株形态特征。随机选择25个花枝,调查花枝数量、花朵数,上下唇长宽。解剖花朵,观察花的内部结构特征。
2.2 开花动态研究
定位观察小陇山林科所院内灰楸的开花、散粉情况,重点观察花朵时空动态,尤其注意柱头与花药的位置,记录花朵开放、花粉散出、柱头伸长、花蜜与气味有无及开始出现和持续的时间等。在花散粉后,注意观察柱头的位置变化。
2.3 杂交指数估算
按照Dafni[3]的标准进行花序直径、花朵大小和开花行为的测量及繁育系统的评判。
2.4 花粉—胚珠比的估算
随机选取灰楸即将开放的花(花药尚未开裂)共30个,每株采10朵,然后细心地解剖花药,将花粉全部移入一个有刻度的离心管,用蒸馏水定容至1 mL。在振荡器下震荡60 s,用移液枪吸取10 μL的花粉液于载玻片上,每朵雄花重复8次,在显微镜下观察并统计花粉数。取30朵雌花,用刀片纵剖子房,记录胚珠数目,按Cruden[4]的标准,花粉—胚珠比为2.7~5.4时,其繁育系统为闭花受精;花粉—胚珠比为18.1~3 910.0时,繁育系统为专性自交;花粉—胚珠比为31.9~39 610.0时,繁育系统为兼性自交;花粉—胚珠比为244.7~2 588.0时,繁育系统为兼性异交;花粉—胚珠比为2 108.0~195 525.0时,繁育系统为专性异交。
2.5 控制授粉实验
用套袋及人工授粉的方法观察花序内、花序间、单花的结实情况。依Dafni[3]描述的方法进行下述处理:(1)对照。不套袋、不去雄、自由传粉,用于检测自然条件下的传粉情况;(2)开花前用细眼沙网套袋、不去雄、自然授粉,检测是否需要传粉者;(3)人工自花授粉,检查是否自交亲合;(4)人工同株异花授粉,检查是否自交亲合;(5)人工异株异花授粉。检查是否杂交亲合。每处理不少于25朵花。
试验花序除自然授粉外,均在花瓣刚刚打开,花药未裂,柱头表面干净无花粉时进行。在试验期间,对套袋的花经常进行检查,并揭袋换气,防止花朵霉坏,揭袋严格防止昆虫在花序内活动。子房明显膨大时,将硫酸纸袋去掉。授粉后15 d调查结实情况。
2.6 数据分析
用Excel 2003及SPSS13.0软件进行数据分析。
3 结果与分析
3.1 花序形态特征
观察表明灰楸花白色到紫红色,花冠筒状,分上下两鄂唇,上鄂唇2裂,下鄂唇3裂(偶见2裂)。上鄂唇长度3.46 cm,变异幅度2.5 ~5.7 cm。下鄂唇长度3.73 cm,变异幅度1.8~5.2 cm。能育雄蕊 2枚,少数3枚。花粉团2,退化雄蕊3枚。雌蕊1枚。是雌雄同花的植物。花冠筒基部有蜜腺。雄蕊与雌蕊均为白色,花丝上有不规则的紫红色斑点,花药乳白色,柱头浅紫红色。雄蕊与雌蕊异熟,雌蕊先熟、雄蕊后熟。对不同居群花数量特征的调查表明,灰楸花朵数最少的1朵,最多的18朵,平均为9.63朵/花枝、小花枝数量平均为9.60个,可见灰楸的花序为总状花序,有时有二歧分枝。
3.2 花期与开花特性
灰楸花期4月底至5月中旬。灰楸展叶后4 d可见花蕾,孕蕾期到开花期约1个月。
依据开花进程,将花期分为5个时期:(1)花蕾期(开花前30 d):又可分为现蕾期,顶芽芽鳞裂开,可见赭黑色花蕾;膨大期:展叶后芽快速生长到(13.19±2.97)cm;绽放期;花蕾赭黑色,密被白色茸毛,膨大成椭圆形,花萼2~3裂。(2)初开期(持续2~3 d):花瓣逐步张开呈钟形,花瓣白色,柱头伸长,已明显大于花药长但低于花冠,但仍呈浅紫红色。花冠筒外黄绿色,上部有不规则紫云斑,花冠筒内基部有紫色纵纹,从花瓣裂口到基部有黄色斑块,基部有蜜腺。花瓣5裂,顶端波纹状,柱头呈浅紫红色,雄蕊呈乳白色,均未开裂。(3)盛开期(持续5~7 d):花瓣完全展开,柱头浅紫红色,打开呈r形,早开的花柱头关闭,颜色加深;花药有一部分开始开裂、散粉。(4)衰落期(持续2~5 d):少部分柱头仍是浅紫红色,多数柱头紫红色,以前离花柱较远的花药完全开裂,花粉逐步散出,花药呈黄色。最早开裂的花药已萎缩,变小,颜色逐渐加深呈深黄色。(5)凋谢期(5~7 d):花粉基本散尽,花药全部萎缩,子房与花药的颜色加深。花瓣仍为白色,但已失去了盛开期的白亮光泽,花瓣和花药一起从花托上脱落下来,见图1。

图1 灰楸开花进程及雌雄配子的发育变化情况
3.3 花粉与柱头可授性检测
观察发现灰楸在花苞刚打开时,雌蕊长度已与花冠筒等长或高于花冠筒,少数雌蕊柱头低于花冠筒,雌蕊不再伸长,第2天柱头打开呈r状,等待授粉。初花时,雄蕊低于1/2花冠筒。花张开后,雄蕊迅速伸长,第2~3天雄蕊伸长靠近雌蕊,花粉团反转90°,与雌蕊平行。水份减少,花粉团逐渐开裂,开始散粉。但柱头多数关闭,由粉色变为褐色,枯萎。之后2 d左右,花粉团干枯。
3.4 有性繁育系统
灰楸杂交指数:按照Dafni[3]的标准方法计算灰楸杂交指数,灰楸花冠口直径为(3.91±0.4675)cm,记为3;花为两性花,雌蕊先熟,记为0;雌雄蕊空间分离,且柱头高于花药,记为1,由此得出灰楸的杂交指数等于4。根据Dafni[3]的标准,该树种的繁育系统为异交、部分自交亲和需要传粉者。
灰楸的花粉—胚珠比:灰楸具雄蕊5个,2强3弱。仅计算2强雄蕊的花粉量,每朵花的花粉量26 750~146 520粒,花粉为四合体,平均为60 222±4 459粒。每朵花有胚珠数为149~256个,平均为195.83±29.26 个。灰楸的花粉—胚珠比为 309,依据Cruden的标准,繁育系统属于兼性异交。
通过套袋及人工授粉的试验,结果表明:灰楸人工自花授粉,结实率为4.08%。灰楸12个同花自交组合有2个结实,但未收到种子(表1)。22个异花自交组合有2个结实,收到种子,但种子全是空粒。表明灰楸自交,不论是同花还是异花自交,均不结实或楸吊在发育过程中败育,这可能是为了避免自交衰退,雌蕊和雄蕊间存在生殖隔离造成的。套袋防虫网袋的组合能结实,表明风是灰楸的传粉媒介之一。灰楸自由授粉的24组合有21个未结实,结实率最高的仅11.11%。范家坑1号自由授粉的结实率最低为0,但是以古灰楸为父本,进行人工授粉结实率为35.897%,说明灰楸自由授粉不结实的原因可能是缺少有效的传粉者,也可能是父本灰楸存在雄性不育或者与母本雌蕊的亲合性低。
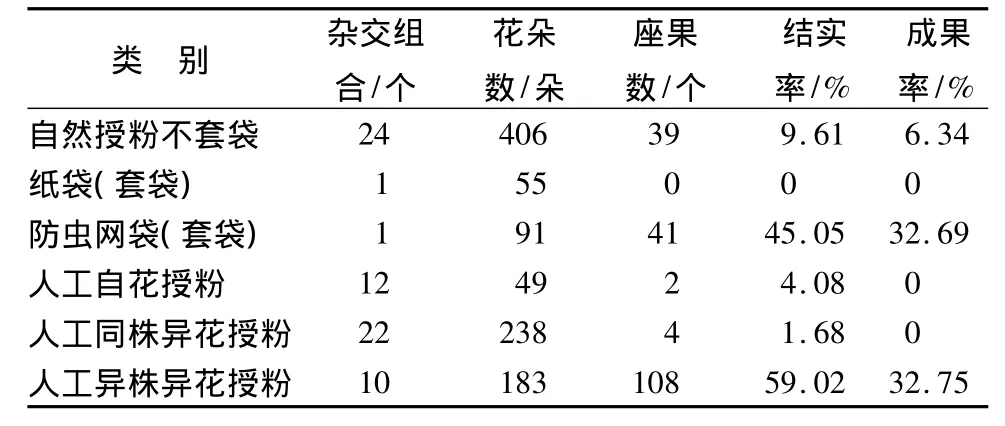
表1 灰楸套袋及人工授粉结果
人工异株异花授粉(不去雄、不套袋)结实率为59.02%。表明人工自花传粉比人工异花传粉的结实率低,反映出该树种较强的异花传粉倾向。在自然条件下,灰楸自由授粉率为9.61%。开花前用细眼沙网套袋、不去雄、自然授粉,结实率为45.05%。两者的结实率均较自交的结实率高、较人工异交的结实率低,表明灰楸自由授粉、细眼沙网套袋授粉均接受了一定的异株花的花粉,两者的传粉者相似,表明风是灰楸传粉者之一。
灰楸的传粉者:观察发现,蜜蜂、马蜂是灰楸的主要访花昆虫,具较高的访花频率,但它在一个花序上停留的时间不长,常为0.5~2 min。访花时间大多集中在11:00—14:00。晴天的访花者数量多,雨天或阴天的访花者数量少。盛花期花序上访花者的访花频率最高。
灰楸同一花序内的筒状花分别位于不同单花时期,而且筒状花花蜜位于筒基部,蜜蜂吸食花蜜的过程中后足或腹部容易接触和附着成熟花粉。当花粉被蜜蜂带走,再访问另一植株的花序时,后足或腹部又容易接触其他花已经展平的二裂柱头,完成灰楸的异花传粉过程。访花者还包括蚂蚁、蝽和蝇等其他昆虫,它们主要活动于花序内部,啃花部幼嫩器官,对灰楸的传粉效果有待于进一步检测。
4 结论与讨论
4.1 灰楸的繁育类型
自然界绝对自交或杂交的植物类群很少,大多是两者兼而有之并采取混和交配的繁殖对策模式[6]。灰楸杂交指数为4,判定繁育系统为异交、部分自交亲和需要传粉者;花粉—胚珠比为309,判定繁育系统为兼性异交;套袋—人工授粉试验结果显示,灰楸为异交为主、部分自交亲合,需要传粉者。3种方法判定的结果完全一致。因此判定,灰楸的繁育系统为兼性异交,以异交为主、部分自交亲合,需要传粉者。人们对楸树的繁育系统认识比较模糊,郭从俭[12]、潘庆凯等[13]认为楸树自花不孕,我们在试验中发现灰楸自花可孕,但自交结实率低。存在自交衰退。
4.2 灰楸的传粉系统
灰楸具有保证异交的许多特征:花序中花朵数目较多、下萼片较大、具两个黄斑、颜色鲜艳、具蜜腺、花粉粒多、柱头面积大并具许多突起、灰楸的柱头与花冠平行或部分在灰楸具有保证异交的许多特征:花序中花朵数目较多、下萼片较大、具两个黄斑、颜色鲜艳、具蜜腺、花粉粒多、柱头面积大并具许多突起、灰楸的柱头部分在花冠外。
对灰楸的传粉媒介检测结果表明,灰楸为风媒、虫媒植物,与传粉媒介的关系属泛化类型。灰楸表现为散粉期滞后于可授期。一般可授期开始较散粉期早1~3 d,结束较散粉期早2~5 d,并且居群散粉期和可授期的重叠期较长,确保在不同时间发育成熟的雌蕊完成授粉、受精作用;另一方面单株花期长,预示着传粉、授粉相对不集中,这可以避免花期恶劣的自然条件(如急骤降温,大雨等)使传粉、受精这一生殖过程中断,这也是灰楸长期适应环境条件所形成的一种生殖策略。
4.3 灰楸的生殖补偿机制
为了保证种群的繁衍和后代的生存,生物往往发展出一整套必要的生殖补偿机制,以保证物种的延续。灰楸自由授粉的结实率低,在有性繁殖受阻的情况下,以根茎进行旺盛的无性繁殖,保持了较高的种群繁衍能力。正常年份,灰楸花期4月底至5月中旬,顶芽分化为花芽,顶芽下部的芽发育为叶芽。如果遇到严重晚霜,灰楸花芽受到冻害,顶部花芽不能正常开花时,顶芽下部的芽将发育为花芽,同时花期延后到5月底到6月中旬。保证了灰楸有性生殖。
不同种群因生境条件的不同,花期、访花者可能不同,交配系统也有可能发生改变。灰楸种群大小、密度和开花数量等影响访花者的访花频率和行为的机制并不清楚,这些问题都有待进一步研究。
[1]潘庆凯,康平生,郭明.楸树[M].北京:中国林业出版社,1991:1-4.
[2]Wyatt R.Pollinator plant interactions and the evolution of breeding systems[M].Orlando:Academy Press,1983:51-95.
[3]Dafni A.Pollinatlon Ecology:a Practical Approach[M].New York:Oxford Univ Press,1992:59-89.
[4]Cruden R W.Pollen ovule ratios:A conservative indicator of breeding system s in flowering plants[J].Evolution,1977,31(1):32-46.
[5]何亚平,刘健全.植物繁育系统研究的最新进展和评述[J].植物生态学报,2003,27(2):151-163.
[6]张金菊,叶其刚,姚小洪,等.片断化生境中濒危植物黄梅秤锤树的开花生物学、繁育系统与生殖成功的因素[J].植物生态学报,2008,32(4):743-750.
[7]李昆,尹伟伦,罗长维.小桐子繁育系统与传粉生态学研究[J].林业科学研究,2007,20(6):775-781.
[8]李鹂,党承林.短葶飞蓬(Erigeron breviscapus)的花部综合特征与繁育系统[J].生态学报,2007,27(2):571-578.
[9]王翠翠,刘文哲,张莹.喜树开花特性及繁育系统的研究[J].热带亚热带植物学报,2009,17(3):275-282.
[10]宋玉霞,郭生虎,牛东玲等.濒危植物肉苁蓉(Cistanche deserticola)繁育系统研究[J].植物研究,2008,28(3):278-288.
[11]Barrett S C H,Harder L D.Ecology and evolution of plant mating[J].Trends in Evolution and Ecology,1996,11(2):73-79.
[12]Worley A C,Baker A M,Thomp son J D,et al.Floral display in Narcissus:variation in flower size and number at the species,population,and individual levels[J].International Journal of Plant Science,2000,161(1):69-79.
猜你喜欢
经济林研究(2022年2期)2022-07-20
课外生活·趣知识(2022年6期)2022-07-14
现代农业科技(2022年4期)2022-03-08
铜仁学院学报(2018年7期)2018-09-08
现代园艺(2017年22期)2018-01-19
中国稻米(2017年6期)2017-12-28
广西植物(2016年10期)2016-11-11
红蜻蜓·低年级(2016年9期)2016-05-14
作物研究(2015年4期)2015-03-24
浙江农业科学(2013年6期)2013-11-08
